Мотокультиватор Крот
Содержимое
- 1 Обзор популярных моделей
- 1.1 Мотокультиватор МК-1А
- 1.2 Мотокультиватор Крот 2 с реверсом
- 2 Руководство по эксплуатации мотокультиватора Крот
- 3 Модернизация модели МК-1А
Производство отечественных мотокультиваторов марки «Крот» было налажено в конце 80-х годов. Первая модель МК-1А была укомплектована двухтактным бензиновым мотором мощностью 2,6 л. с. Пуск осуществлялся от веревочного ручного стартера. Изначально техника предназначалась для обработки небольших огородов на даче и работы внутри теплицы. Современный мотокультиватор Крот представляет усовершенствованную модель МК-1А. Такая техника уже комплектуется мощным двигателем с принудительным воздушным охлаждением.
Обзор популярных моделей
Примерные размеры техники находятся в пределах:
- длина – от 100 до 130 см;
- ширина – от 35 до 81 см;
- высота – от 71 до 106 см.
Габариты мотокультиватора Крот зависят от модели, и с усовершенствованием техники могут изменяться.
Мотокультиватор МК-1А
Обзор мотокультиваторов Крот начнем с модели МК-1А. Агрегат укомплектован двухтактным карбюраторным мотором мощностью 2,6 л.с. В качестве стартера используется заводная рукоятка на веревке. Бензиновый двигатель с редуктором имеет простейшее болтовое соединение с рамой. Топливный бак рассчитан на 1,8 л. Такой маленький объем обусловлен небольшим расходом топлива. Заправку агрегата можно осуществлять дешевым бензином марки АИ-80 или А-76. Для приготовления топливной смеси применяется машинное масло марки М-12ТП. Весит мотокультиватор всего 48 кг. Такую технику легко транспортировать на дачу легковым автомобилем.
Все элементы управления мотокультиватора расположены на рукоятках, а именно:
- рычаг сцепления;
- рычаг управления дроссельной заслонки;
- рычаг управления карбюраторной заслонки.
Модель Крот MK-1А способна работать с навесным оборудованием. Мотокультиватор используют для поливки, покоса травы, обработки грунта и ухода за насаждениями.
Мотокультиватор Крот 2 с реверсом
Особенностью конструкции является то, что обладает мотокультиватор Крот реверсом и мощным двигателем. Это дает возможность потребителю получить за малые деньги настоящий мотоблок. Агрегат работает от четырехтактного бензинового мотора Honda GX200 мощностью 6,5 л. с. Крот 2 имеет электронное зажигание, вал отбора мощности, бак для бензина емкостью 3,6 л. Крутящий момент от мотора к ходовой части передается ременной передачей.
Среди других мотокультваторов с подобными характеристиками, эта модель Крота по надежности занимает первые позиции. Таких показателей удалось добиться благодаря мощному одноцилиндровому мотору и безотказному редуктору. Моторесурс двигателя составляет 3500 часов. Это довольно много по сравнению со старыми моделями мотокультиватора Крот, у которых моторесурс доходил до 400 часов.
Важно! Большой плюс четырехтактного двигателя в том, что масло с бензином находятся отдельно. Хозяину теперь не нужно вручную готовить топливную смесь, смешивая эти компоненты.
Мощности мотокультиватора с задним ходом хватает для захвата фрезами участка шириной 1 м. Инструкция по эксплуатации от завода производителя гласит, что мотокультиватор Крот 2 способен расширять свою функциональность за счет использования навесного оборудования. Так, техника может стать снегоуборщиком или косилкой, транспортом для перевозки груза, машиной для выполнения многих сельскохозяйственных работ.
Важно! Ручки мотокультиватора Крот 2 обладают многоступенчатой регулировкой. Оператор может повернуть их в любое направление, что дает возможность оптимально подстроить технику для любых типов работ.
На видео предлагаем посмотреть обзор мотокультиватора Крот:
Руководство по эксплуатации мотокультиватора Крот
Итак, мы выяснили, что современный мотокультиватор Крот обладает почти всеми функциями мотоблока. Сейчас давайте ознакомимся, что гласит руководство по эксплуатации рассматриваемой техники:
- Прямое назначение мотокультиватора – это вспашка земли.
 Делается это при помощи фрез, которые насаживают на валы редуктора. Транспортные колеса при вспашке поднимают. Сзади к прицепной скобе закреплен сошник. Он используется как тормоз, а также для регулировки глубины обработки грунта. Двигается мотокультиватор за счет вращения фрез, при этом одновременно происходит рыхление почвы. В комплекте с агрегатом идут две внутренние и наружные фрезы. Первый тип применяют на грубом грунте и целине. Легкую землю рыхлят обеими фрезами, а можно добавить и третий комплект. Его покупают отдельно. В итоге получается с каждой стороны по три фрезы, а в сумме их 6 штук. Восемь фрез на мотокультиватор Крот ставить нельзя из-за возрастания нагрузки на мотор и трансмиссию.
Делается это при помощи фрез, которые насаживают на валы редуктора. Транспортные колеса при вспашке поднимают. Сзади к прицепной скобе закреплен сошник. Он используется как тормоз, а также для регулировки глубины обработки грунта. Двигается мотокультиватор за счет вращения фрез, при этом одновременно происходит рыхление почвы. В комплекте с агрегатом идут две внутренние и наружные фрезы. Первый тип применяют на грубом грунте и целине. Легкую землю рыхлят обеими фрезами, а можно добавить и третий комплект. Его покупают отдельно. В итоге получается с каждой стороны по три фрезы, а в сумме их 6 штук. Восемь фрез на мотокультиватор Крот ставить нельзя из-за возрастания нагрузки на мотор и трансмиссию. - При прополке сорняков выполняют переоборудование механизма. На внутренних фрезах снимают ножи, а вместо них ставят полольники. Эти детали узнаются по Г-образной форме. Внешние фрезы заменяют дисками. Они тоже покупаются отдельно. Диски нужны для защиты растений, препятствуя попаданию их под полольник. Если прополка осуществляется по картофелю, то одновременно можно выполнять предварительное окучивание. Для этого установленный сзади сошник заменяют окучивателем.
- Когда требуется окучить картофель, фрезы не нужны. Их снимают с вала редуктора, а на это место ставят стальные колеса с наваренными грунтозацепами. Окучиватель остается на своем месте, где раньше стоял сошник.
- Во время уборки картофеля используют те же металлические грунтозацепы, а сзади мотокультиватора сошник заменяют картофелекопалкой. Этот тип навесного оборудования выпускается разных модификаций, но к культиваторам обычно покупают веерные модели.
- Вспашку земли можно делать не только фрезами, но и плугом. Его крепят сзади агрегата на место сошника. Стальные колеса остаются на месте.
- Использовать агрегат можно при сенокосе. Нужно лишь купить косилку и закрепить ее спереди агрегата. На валы редуктора надевают резиновые колеса. Передачу крутящего момента обеспечивают ремни, надетые на шкивы мотокультиватора Крот и косилки.
- Крот отлично способен заменить помпу для перекачивания воды. Нужно лишь приобрести насосное оборудование МНУ-2, закрепить его на раме и соединить ременной передачей. При этом важно не забыть снять ремень с тягового редуктора.
- Мотокультиватор неплохо справляется с перевозкой малогабаритных грузов весом до 200 кг. Здесь потребуется тележка с поворотно-сцепным механизмом. Можно приобрести модельТМ-200 заводского изготовления или сварить самостоятельно из металла. Во время транспортировки грузов, на валы редуктора надевают резиновые колеса.
 Делается это при помощи фрез, которые насаживают на валы редуктора. Транспортные колеса при вспашке поднимают. Сзади к прицепной скобе закреплен сошник. Он используется как тормоз, а также для регулировки глубины обработки грунта. Двигается мотокультиватор за счет вращения фрез, при этом одновременно происходит рыхление почвы. В комплекте с агрегатом идут две внутренние и наружные фрезы. Первый тип применяют на грубом грунте и целине. Легкую землю рыхлят обеими фрезами, а можно добавить и третий комплект. Его покупают отдельно. В итоге получается с каждой стороны по три фрезы, а в сумме их 6 штук. Восемь фрез на мотокультиватор Крот ставить нельзя из-за возрастания нагрузки на мотор и трансмиссию.
Делается это при помощи фрез, которые насаживают на валы редуктора. Транспортные колеса при вспашке поднимают. Сзади к прицепной скобе закреплен сошник. Он используется как тормоз, а также для регулировки глубины обработки грунта. Двигается мотокультиватор за счет вращения фрез, при этом одновременно происходит рыхление почвы. В комплекте с агрегатом идут две внутренние и наружные фрезы. Первый тип применяют на грубом грунте и целине. Легкую землю рыхлят обеими фрезами, а можно добавить и третий комплект. Его покупают отдельно. В итоге получается с каждой стороны по три фрезы, а в сумме их 6 штук. Восемь фрез на мотокультиватор Крот ставить нельзя из-за возрастания нагрузки на мотор и трансмиссию. Если прополка осуществляется по картофелю, то одновременно можно выполнять предварительное окучивание. Для этого установленный сзади сошник заменяют окучивателем.
Если прополка осуществляется по картофелю, то одновременно можно выполнять предварительное окучивание. Для этого установленный сзади сошник заменяют окучивателем.
Как видите, благодаря дополнительному оборудованию многофункциональность Крота существенно расширяется.
Модернизация модели МК-1А
Если у вас имеется старая модель Крота, не стоит спешить ее выбрасывать. Зачем потом переплачивать при покупке нового культиватора за раму, редуктор и другие детали, если они уже есть. Можно обойтись простой заменой мотора.
Старый двигатель можно заменить на четырехтактный LIFAN — 160F. Китайский мотор не дорогой, плюс обладает мощностью 4 л. с. По паспорту мотокультиватору МК-1А при обработке грунта фрезами на глубину 20 см нужно добавлять обороты. С новым мотором этого делать не надо. Еще с увеличением мощности двигателя изменилась глубина обработки, и теперь она достигает 30 см. На большую глубину рассчитывать не стоит, так как начнется пробуксовка ремня.
Китайский мотор не дорогой, плюс обладает мощностью 4 л. с. По паспорту мотокультиватору МК-1А при обработке грунта фрезами на глубину 20 см нужно добавлять обороты. С новым мотором этого делать не надо. Еще с увеличением мощности двигателя изменилась глубина обработки, и теперь она достигает 30 см. На большую глубину рассчитывать не стоит, так как начнется пробуксовка ремня.
Установить на старую раму новый мотор не сложно. Все крепления практически подходят. Единственная сложность – потребуется переделка родного шкива. Его снимают со старого мотора, рассверливают внутреннее отверстие под диаметр вала нового двигателя, после чего насаживают с применением шпонки.
Если при снятии шкива он случайно треснул, не спешите бежать за новым. Можно попробовать отреставрировать его с помощью холодной сварки. Как это сделать, лучше расскажут на видео:
Крот считается не плохой техникой для маленького участка, но и требовать от него выполнения сверхсложных задач не стоит. Для этих целей существуют тяжелые мотоблоки и мини-трактора.
Для этих целей существуют тяжелые мотоблоки и мини-трактора.
Мотокультиватор Крот МК-1А — технические характеристики, устройство
Благодаря высокой в своё время популярности мотокультиватор Крот МК-1А обзавёлся парой модификаций, которые несколько отличаются своими характеристиками, сохраняя всё же сходство с оригиналом. При этом сходство заключается не только в качестве работы, но и во внешних признаках. Встретить его можно где угодно, т.к. выпускается он с начала 80-х прошлого века.
Мотокультиватор Крот МК-1А в работе
Устройство мотокультиватора Крот МК-1А
По сравнению со своим предшественником мотокультиватор Крот МК-1А новой модификации имеет более эргономичную форму рукоятей, а также больший вес, что увеличивает проходимость при первичной обработке грунта. Помимо основного своего назначения — вспашки почвы, он со значительным успехом справляется с рядом других задач, среди которых числятся:
- Прополка сорняков;
- Окучивание грядок;
- Выкапывание картофеля;
- Сенокос;
- Перекачка воды;
- Перевозка небольших грузов.

Однако для выполнения данных операций потребуется приобретать дополнительно комплект навесного оборудования, состоящий из таких элементов:
- полольников;
- окучивателя;
- колёс с грунтозацепами;
- выкапывателя;
- плуга;
- косилки;
- насосной установки;
- тележки.
Такая оптимизация позволяет классифицировать инструмент не как мотокультиватор, а как мотоблок.
В зависимости от комплектации он может иметь 2-х или 4-хтактный двигатель. При использовании первого увеличивается моторесурс изделия, а во втором случае повышается силовая тяга, что гораздо удобнее на сложно проходимых участках. Мотор у мотокультиватора имеет воздушное охлаждение. Переключение передач у него – механическое, однако, в некоторых версиях движение может осуществляться только вперёд. Самый облегчённый вариант мотокультиватора Крот МК-1А имеет вес – 48 кг, что обеспечивает ему отличную мобильность в междурядьях.
Внешний вид мотокультиватора Крот МК-1А
Расхода топлива культиватора Крот МК-1А
Экономичный расход топлива обеспечивается за счёт использования центробежного регулятора частоты вращения коленчатого вала, подключённого к двигателю. Также этому способствует переработанный карбюратор и воздушный фильтр, а также реверсивный режим работы, обеспечивающий задний ход. В целом расход топлива не превышает 1 литра за час работы. В качестве основного вида топлива применяется низкооктановый бензин марки А-76 в смеси с маслом МГ-8А или М-12 ТП. Для удобства работы ёмкость топливного бака ограничивается объёмом 1,8 литра.
Также этому способствует переработанный карбюратор и воздушный фильтр, а также реверсивный режим работы, обеспечивающий задний ход. В целом расход топлива не превышает 1 литра за час работы. В качестве основного вида топлива применяется низкооктановый бензин марки А-76 в смеси с маслом МГ-8А или М-12 ТП. Для удобства работы ёмкость топливного бака ограничивается объёмом 1,8 литра.
О недостатках мотоблока
Многофункциональный мотоблок Крот имеет свои недостатки:
Мотоблок Крот в работе
- Основной недостаток устройства – маломощный двигатель. При большем объеме возможностей стало бы больше.
- Конструкция открытого типа приводит к сильному загрязнению при работе. Пылью забиваются труднодоступные места, в которых грязи быть не должно.
- В промежуток между корпусом и ножами попадают камни, корни, комья земли, затрудняющие работу.
- Только одна рабочая скорость.
- Ресурс работы 2-хтактного двигателя не превышает 400 моточасов.
- За один проход обрабатывается узкая полоса земли.
- Поршневая группа через 1-2 года выходит из строя и подлежит замене.
- После замены двигателя на модель мощнее, раму приходится укреплять дополнительными деталями.
- Высокая цена агрегата.
 При большем объеме возможностей стало бы больше.
При большем объеме возможностей стало бы больше.Технические характеристики культиватора Крот МК-1А
- Двухтактный карбюраторный двигатель с воздушным охлаждением;
- мощность двигателя при 5500-6500 об. /мин. — 1,9 кВт, 2.6 л.с.; (зависит от модели)
- объем топливного бака — 1,8 л;(зависит от медели)
- ширина обработки — 350, 600 мм;
- одна передача – вперед;
- масса – 48 кг.;(зависит от модели)
- габаритные размеры в рабочем положении 1300x810x1060 мм.;
- глубина обработки — до 250 мм.;
- производительность при фрезеровании — 150-200 м2/час.
 /мин. — 1,9 кВт, 2.6 л.с.; (зависит от модели)
/мин. — 1,9 кВт, 2.6 л.с.; (зависит от модели)Задняя скорость в домашних условиях
Задняя скорость на мотоблоке “Крот” присутствуют далеко не во всех моделях. Иногда это зависит даже от того, где агрегат был произведен – в Москве или Омске. Благо, умельцы придумали несложный способ улучшить мотоблок “Крот”. Модернизации бывают разными.
Любителям самостоятельных доработок возможно заинтересует самодельная зернодробилка.
Вот один из рецептов, как сделать заднюю скорость на мотоблоке “Крот”. Откручивается гайка, которая держит шкив сельскохозяйственного аппарата. Нанизываем рядом с ним шкив от, к примеру, “Волги”. Гайку прикручиваем.
Гайку прикручиваем.
В конструкции прижимного ролика используем ремень ГРМ от любой иномарки. И, собственно, переключатель в передней части мотоблока – небольшая металлическая планка, соединенная со всем механизмом прижимного ролика. Планка крепится на двух пружинах и от нее идет тросик к ручке переключателя на рукояти.
Мотокультиватор Крот МК 1а: инструкция по эксплуатации
Производство отечественных мотокультиваторов марки «Крот» было налажено в конце 80-х годов. Первая модель МК-1А была укомплектована двухтактным бензиновым мотором мощностью 2,6 л. с. Пуск осуществлялся от веревочного ручного стартера. Изначально техника предназначалась для обработки небольших огородов на даче и работы внутри теплицы. Современный мотокультиватор Крот представляет усовершенствованную модель МК-1А. Такая техника уже комплектуется мощным двигателем с принудительным воздушным охлаждением.
Руководство по эксплуатации мотокультиватора Крот
Итак, мы выяснили, что современный мотокультиватор Крот обладает почти всеми функциями мотоблока. Сейчас давайте ознакомимся, что гласит руководство по эксплуатации рассматриваемой техники:
Сейчас давайте ознакомимся, что гласит руководство по эксплуатации рассматриваемой техники:
- Прямое назначение мотокультиватора – это вспашка земли. Делается это при помощи фрез, которые насаживают на валы редуктора. Транспортные колеса при вспашке поднимают. Сзади к прицепной скобе закреплен сошник. Он используется как тормоз, а также для регулировки глубины обработки грунта. Двигается мотокультиватор за счет вращения фрез, при этом одновременно происходит рыхление почвы. В комплекте с агрегатом идут две внутренние и наружные фрезы. Первый тип применяют на грубом грунте и целине. Легкую землю рыхлят обеими фрезами, а можно добавить и третий комплект. Его покупают отдельно. В итоге получается с каждой стороны по три фрезы, а в сумме их 6 штук. Восемь фрез на мотокультиватор Крот ставить нельзя из-за возрастания нагрузки на мотор и трансмиссию.
- При прополке сорняков выполняют переоборудование механизма. На внутренних фрезах снимают ножи, а вместо них ставят полольники. Эти детали узнаются по Г-образной форме. Внешние фрезы заменяют дисками. Они тоже покупаются отдельно. Диски нужны для защиты растений, препятствуя попаданию их под полольник. Если прополка осуществляется по картофелю, то одновременно можно выполнять предварительное окучивание. Для этого установленный сзади сошник заменяют окучивателем.
- Когда требуется окучить картофель, фрезы не нужны. Их снимают с вала редуктора, а на это место ставят стальные колеса с наваренными грунтозацепами. Окучиватель остается на своем месте, где раньше стоял сошник.
- Во время уборки картофеля используют те же металлические грунтозацепы, а сзади мотокультиватора сошник заменяют картофелекопалкой. Этот тип навесного оборудования выпускается разных модификаций, но к культиваторам обычно покупают веерные модели.
- Вспашку земли можно делать не только фрезами, но и плугом. Его крепят сзади агрегата на место сошника. Стальные колеса остаются на месте.
- Использовать агрегат можно при сенокосе. Нужно лишь купить косилку и закрепить ее спереди агрегата. На валы редуктора надевают резиновые колеса. Передачу крутящего момента обеспечивают ремни, надетые на шкивы мотокультиватора Крот и косилки.
- Крот отлично способен заменить помпу для перекачивания воды. Нужно лишь приобрести насосное оборудование МНУ-2, закрепить его на раме и соединить ременной передачей. При этом важно не забыть снять ремень с тягового редуктора.
- Мотокультиватор неплохо справляется с перевозкой малогабаритных грузов весом до 200 кг. Здесь потребуется тележка с поворотно-сцепным механизмом. Можно приобрести модельТМ-200 заводского изготовления или сварить самостоятельно из металла. Во время транспортировки грузов, на валы редуктора надевают резиновые колеса.
 Эти детали узнаются по Г-образной форме. Внешние фрезы заменяют дисками. Они тоже покупаются отдельно. Диски нужны для защиты растений, препятствуя попаданию их под полольник. Если прополка осуществляется по картофелю, то одновременно можно выполнять предварительное окучивание. Для этого установленный сзади сошник заменяют окучивателем.
Эти детали узнаются по Г-образной форме. Внешние фрезы заменяют дисками. Они тоже покупаются отдельно. Диски нужны для защиты растений, препятствуя попаданию их под полольник. Если прополка осуществляется по картофелю, то одновременно можно выполнять предварительное окучивание. Для этого установленный сзади сошник заменяют окучивателем. Нужно лишь купить косилку и закрепить ее спереди агрегата. На валы редуктора надевают резиновые колеса. Передачу крутящего момента обеспечивают ремни, надетые на шкивы мотокультиватора Крот и косилки.
Нужно лишь купить косилку и закрепить ее спереди агрегата. На валы редуктора надевают резиновые колеса. Передачу крутящего момента обеспечивают ремни, надетые на шкивы мотокультиватора Крот и косилки.Обзор навесного оборудования
Навесное оборудование используется для улучшения производительности мотоблока, делая его универсальным средством. Перечень дополнительного оборудования довольно широкий:
- Фрезы. Саблевидные – могут быть 4 или 6 фрез и из треугольных наконечников – разрезают любую поверхность земли без налипания.
- Адаптер. Сиденье, рама, дополнительные колеса, установленные на мотоблок. В такой комплектации крот мотоблок похож на мини трактор.
- Прицеп. Можно оснастить мотоблок прицепной тележкой разных габаритов и грузоподъемности. Может переводить до 500 кг веса. Используется при транспортировке сыпучих грузов.
- Косилки. С помощью этого дополнительного оборудования можно использовать мотоблок как сенокосилку.
- Колеса. Обязательны для перевозки конструкции, идут в комплекте у всех моделей Крота. Колеса имеют мощные протекторы, что улучшает сцепление с землей и проходимость агрегата.
- Грунтозацепы. Вместо пневматических колес можно использовать грунтозацепы. Сделаны из металла, с наваренными ребрами, вспушивания земли ребрами.
- Плуг. Используется для перепахивания земли и борьбы с сорняками.
- Снегоуборщик. Использовав этот навесной блок Крот можно использовать в качестве снегоуборочной машины. Насадки для уборки снега бывают двух видов – шнековый снегоотбрасыватель и щетка.
- Картофелесажалка. С помощью этого оборудования можно механизировать посадку картофеля. Как это работает? Засыпается картофель для посадки, устройством захватывается одна картофелина и укладывается в подготовленную до этого борозду на нужную глубину. Сверху формируется холмик земли. Заранее настраивается расстояние между клубнями, и процесс пошел.
- Картофелекопатель. С помощью этого оборудования можно легко и быстро собрать посаженный урожай. Картофель аккуратно захватывается, вынимается, земля просеивается, а овощ остается на специальных прутьях.
- Утяжелитель. Необходим чтобы оборудование могло глубже погрузиться в землю. Чаще всего используется на легких агрегатах с весом до 100 кг.
- Сцепка. Является одним из важнейших частей мотоблока. Способствует соединению мотоблока с любым навесным дополнительным оборудованием.
 Саблевидные – могут быть 4 или 6 фрез и из треугольных наконечников – разрезают любую поверхность земли без налипания.
Саблевидные – могут быть 4 или 6 фрез и из треугольных наконечников – разрезают любую поверхность земли без налипания. Насадки для уборки снега бывают двух видов – шнековый снегоотбрасыватель и щетка.
Насадки для уборки снега бывают двух видов – шнековый снегоотбрасыватель и щетка.
При помощи оборудования можно расширить сферу применения мотоблока
Модернизация модели МК-1А
Если у вас имеется старая модель Крота, не стоит спешить ее выбрасывать. Зачем потом переплачивать при покупке нового культиватора за раму, редуктор и другие детали, если они уже есть. Можно обойтись простой заменой мотора.
Зачем потом переплачивать при покупке нового культиватора за раму, редуктор и другие детали, если они уже есть. Можно обойтись простой заменой мотора.
Старый двигатель можно заменить на четырехтактный LIFAN — 160F. Китайский мотор не дорогой, плюс обладает мощностью 4 л. с. По паспорту мотокультиватору МК-1А при обработке грунта фрезами на глубину 20 см нужно добавлять обороты. С новым мотором этого делать не надо. Еще с увеличением мощности двигателя изменилась глубина обработки, и теперь она достигает 30 см. На большую глубину рассчитывать не стоит, так как начнется пробуксовка ремня.
Установить на старую раму новый мотор не сложно. Все крепления практически подходят. Единственная сложность – потребуется переделка родного шкива. Его снимают со старого мотора, рассверливают внутреннее отверстие под диаметр вала нового двигателя, после чего насаживают с применением шпонки.
Если при снятии шкива он случайно треснул, не спешите бежать за новым. Можно попробовать отреставрировать его с помощью холодной сварки. Как это сделать, лучше расскажут на видео:
Как это сделать, лучше расскажут на видео:
Крот считается не плохой техникой для маленького участка, но и требовать от него выполнения сверхсложных задач не стоит. Для этих целей существуют тяжелые мотоблоки и мини-трактора.
Возможные неисправности и ремонт
В процессе эксплуатации мотоблока могут возникать разные поломки и сбои. Из-за этого у фермеров появляется желание отремонтировать агрегат Крот своими руками. В большинстве случаев проблемы возникают с такими рабочими органами:
- Двигатель.
- Магнето, система зажигания.
- Редуктор.
- Воздушные фильтры.
- Карбюратор.
Чтобы понять, как осуществить ремонт культиватора Крот, следует подробно разобраться с основными причинами неполадок и их решением.
Как завести
Если мотокультиватор Крот не заводится, возможно это связано со следующими поломками и проблемами:
- Отсутствие искры. Возможно произошло перегорание искры зажигания. Для устранения дефекта нужно заменить свечу.
- Свеча покрылась копотью. Чтобы восстановить ее работоспособность, достаточно выполнить глубокую чистку бензином.
- При наличии хорошей искры, но отсутствии запуска двигателя, следует посмотреть на качество изоляции. Порой возникает необходимость замены наконечника.
- Низкое качество топливной смеси.
- Стекание топлива со свечи.
- Проблемы с воздушным фильтром. Если он покрылся загрязнениями, проблем с зажиганием не избежать.
- Повреждение магнето. Подобный узел нельзя восстановить, поэтому понадобится покупка нового.
Как заменить двигатель
Необходимость замены двигателя возникает при его выходе из строя. Проблема часто связана с отсутствием или низким качеством топливной смеси, неполадках в системе зажигания или декомпрессии, вызвавшей износ поршня и деформацию выпускного клапана.
Если неприятность объясняется некачественным топливом, то достаточно заменить его. При наличии более опасных поломок придется выполнить установку двигателя Хонда, Лифан, Субару или другой модели.
Установить движок самостоятельно несложно. Главное — придерживаться схемы, где указаны требуемые параметры монтажа.
Как выставить зажигание
В случае неисправностей в системе зажигания необходимо проверить работоспособность свечи, предварительно выкрутив ее и осмотрев.
Если приспособление сухое, значит топливная смесь не достигает цилиндров.
Мокрая свеча отображает важность просушки цилиндров с помощью ручного стартера. Исправность свечи проверяется мультиметром, который определяет показатели напряжения в ОМ.
Магнето
В случае выхода из строя магнето восстановить его будет невозможно. Единственным решением станет покупка нового исправного устройства.
Редуктор
Роль редуктора заключается в передаче вращательного момента от мотора к валу. Если он выйдет из строя, работа мотоблока окажется невозможной. Чтобы восстановить систему, необходимо разобраться с ее устройством и спецификой ремонта.
Если он выйдет из строя, работа мотоблока окажется невозможной. Чтобы восстановить систему, необходимо разобраться с ее устройством и спецификой ремонта.
Задний ход
Современные модели культиваторов Крот оснащаются опцией реверса (задней передачи), которая расширяет их функциональность и эксплуатационные свойства. Выход из строя реверсной передачи случается редко, но если это произойдет, понадобится заменить ее.
Замена ремня
Заменяя ремень, необходимо определить его размер и рабочие свойства. Их описание указывается в технической документации к мотокультиватору.
Мотокультиватор «Крот»-1а
В конце 1980-х годов прошлого столетия на российских машиностроительных заводах стали производить конструкционно легкие и простые мотокультиваторы «Крот»-1А-02. Первые модели МК-1А-02 были оснащены двухтактным двигателем, его мощность достигала 2,6 лошадиных сил. Также этот агрегат имел редуктор, прикрепленный болтиками к раме. На электромотор закреплялся веревочный не съемный стартер. Агрономы отмечали, что устройство обладало небольшой производительностью электродвигателя. После таких отзывов производители решили существенно усовершенствовать агромашины, в частности, увеличив мощность агрегатов.
Агрономы отмечали, что устройство обладало небольшой производительностью электродвигателя. После таких отзывов производители решили существенно усовершенствовать агромашины, в частности, увеличив мощность агрегатов.
Отзывы владельцев
Что же говорят об этой агротехнике наши эксперты — обычные владельцы дачных участков и огородов, которым посчастливилось купить мотокультиватор “Крот”? Давайте ознакомимся с несколькими отзывами:
Иван Андреевич, 54 года
«В целом, мотокультиватор МК-1А-02 “Крот” мне очень нравится! Пользуюсь этой машиной уже 17 лет, агрегат зарекомендовал себя как надежная техника. За все время эксплуатации было несколько серьезных поломок: накрылся магнето — пришлось заменить, первое время глох движок, но я быстро усек в чем дело — тщательно прочистил карбюратор и впредь с неочищенным от пыли, грязи мотоблоком на работу не выходил. Пару раз приходилось в процессе вспашки поля подтягивать крепежные болты, машина сразу прекращала тарахтеть, как трещетка. Ну, что еще — сальники менял пару раз. Функционал впечатляющий, пришлось подкупить кое-какое оборудование, некоторое сделал сам (снегоуборщик). Мужики, рекомендую всем — не пожалеете!»
Ну, что еще — сальники менял пару раз. Функционал впечатляющий, пришлось подкупить кое-какое оборудование, некоторое сделал сам (снегоуборщик). Мужики, рекомендую всем — не пожалеете!»
Василий, 39 лет
«Первый мотокультиватор Крот был еще у моего отца. Так что при покупке я уже наперед понимал, что покупаю. Меня все устраивает: и очень доступная цена, и хорошая производительность. Читал отзывы, что мол мотоблок для целины не годится — слабоват, отвечу — если пройтись несколькими этапами, да в разных направлениях — подчинится и целина! Купили с женой дом с большим участком, земля такая — думали ничего не получится! Но мотокультиватор приятно удивил, медленно, потихонечку взяли мы эту целину. Здесь главное знать как и не отступать!»
Основные технические спецхарактеристики
«Крот» 1 мотокультиватор оснащается карбюраторным 2-тактным электродвигателем мощностью 2,6 лошадиных сил с воздушной охлаждающей системой и редуктором. И электродвигатель, и редуктор зафиксированы болтами к раме. Мотор на культиватор «Крот» MK-1А становится бензиновый. На электромоторе фиксируется веревочный не съемный стартер. Данная агромашина также оснащена топливным бачком объемом 1,8 литра. Мотокультиватор «Крот» «Крот»-1А успешно работает на бензине АИ-80 или А-76 (с маслом М-12ТП). Масса агромашины составляет 48 килограмм.
Мотор на культиватор «Крот» MK-1А становится бензиновый. На электромоторе фиксируется веревочный не съемный стартер. Данная агромашина также оснащена топливным бачком объемом 1,8 литра. Мотокультиватор «Крот» «Крот»-1А успешно работает на бензине АИ-80 или А-76 (с маслом М-12ТП). Масса агромашины составляет 48 килограмм.
Еще одна важная техническая характеристика — это руль. Он оснащен первоклассными элементами управления, в частности:
- pучками сцeплeния и pучками для проведения pегулиpования дpоссельной заслонки;
- управляющим рычагом карбюраторной заслонки.
Мотокультиватор «Крот» MK-1А обладает навесными конструкциями. Их можно приспособить для окучивания, пропалывания, поливочных работ, скашивания травы.
Как произвести замену
Безотказная работа минитрактора будет зависеть от соблюдения последовательности установки и подключения.
- Открутить заводское крепление и снять кожух.
- Отключить провода и отсоединить ремень.
- Провести демонтаж.
- Приставить новый двигатель и подметить новые крепежные отверстия на раме.
- Установить двигатель и накинуть ремень.
- Отрегулировать натяжение ремня перемещением мотора.
- Выставить агрегат так, чтобы шкив мотора и редуктора были в одной плоскости и располагались соосно.
- Просверлить сквозные отверстия и закрепить двигатель мотоблока болтами длиной 35 мм.
- Подключить сцепление, газ и систему глушителя.
- Провести обкатку на холостых оборотах.

Мотокультиватор Крот МК-1А — технические характеристики, устройство
Содержание статьи
Мотокультиваторы Крот МК-1А-02. Описание модели. Технические характеристики. Особенности эксплуатации
Описание
Вот уже более 35 лет на машиностроительных заводах в Омске и Москве выпускается ветеран полей, садов и огородов мотокультиватор Крот. За это время культиватор подвергался модернизации и различным усовершенствованиям, однако общая конструкция осталась практически неизменной. В настоящее время на культиваторы устанавливаются современные двигатели от известных производителей GeoTeck, Subaru, HAMMERMANN.
В настоящее время на культиваторы устанавливаются современные двигатели от известных производителей GeoTeck, Subaru, HAMMERMANN.
Мотокультиватор Крот МК-1А-02 предназначен для первичной обработки почвы, а также для прополки сорняков, окучивания рядков и грядок, транспортирования грузов, покоса травы, выкапывания картофеля, перекачки воды. Крот МК-1А-02 оснащен двухтактным двигателем мощностью 2,6 л.с. с увеличенным моторесурсом. В зависимости от количества установленных фрез способен обрабатывать грунт с шириной захвата 35-60 см. глубиной до 25 см.
По сравнению с базовой моделью МК-1А, современная версия МК-1А-02 выгодно отличается улучшенными техническими характеристиками и расширенным функционалом:
- Благодаря увеличенной мощности двигателя, возросла производительность агрегата.
- Небольшая масса позволяет легко управлять культиватором человеку даже с небольшой физической силой.
- Агрегат можно удобно транспортировать в багажнике авто.
- Доступная агрегация с различными дополнительными приспособлениями – оригинальными и других производителей.

Технические характеристики модели МК-1А-02
- Двухтактный карбюраторный двигатель с воздушным охлаждением;
- Мощность двигателя при 5500-6500 об./мин. — 1,9 кВт, 2.6 л.с.;
- Топливо — смесь бензина А-76, АИ-80 с маслом М-12ТП;
- Объем топливного бака — 1,8 л;
- Ширина обработки — 350, 600 мм;
- Одна передача — вперед;
- Масса — 48 кг.;
- Габаритные размеры в рабочем положении 1300x810x1060 мм.;
- Глубина обработки — до 250 мм.;
- Производительность при фрезеровании — 150-200 м 2 /час.
Инструкция по эксплуатации: особенности обслуживания
Мотокультиватор Крот МК-1А-02 прост в обслуживании и эксплуатации. Потребляет в качестве горючего смесь бензина АИ-76, АИ-80 с моторным маслом М-12-ТПТУ в соотношении 25:1. Для редуктора двигателя используется масло МГ-8А, для выходного редуктора ТАД-17.
Для нового культиватора первые 15 часов эксплуатации являются периодом обкатки – приработки основных узлов и механизмов.
 В этот период запрещается использовать технику на полную мощность.
В этот период запрещается использовать технику на полную мощность.На мотокультиватор доступно устанавливаются различные навесные орудия: окучники, пропольники, тележка, косилка, мотопомпа, плуг, снегоуборщик. Благодаря заднему ходу, агрегат обладает хорошей маневренностью, особенно на ограниченных пространствах в теплицах, на виноградниках.
Возможные неисправности, их устранение, ремонт
Каждый вид техники отличается определенными особенностями. Это относится и к мотокультиваторам Крот, поэтому неукоснительное соблюдение рекомендаций завода-изготовителя является залогом продолжительной бесперебойной работы машин.
Как свидетельствует опыт большинства владельцев техники, основные причины неполадок и поломок мотокультиватора Крот МК-1А-02 сводятся к одной: загрязнение деталей, узлов и механизмов. Поэтому поддержание сельхозмашины в чистоте и своевременное техническое обслуживание должно быть главным правилом успешной эксплуатации.
- При загрязненном карбюраторе мотокультиватор Крот МК-1А-02 будет перегреваться и глохнуть.
- Двигатель может не развивать достаточной мощности из-за засорения карбюратора, появления нагара в глушителе, на каналах цилиндра, засорения воздушного фильтра двигателя. Причинами также может быть увеличение натяжения клинового ремня, отсутствие компрессии.
- Не используйте в качестве топлива чистый, не смешанный с маслом бензин.
- Нельзя применять моторное масло тех марок, которые не указаны в инструкции по эксплуатации.
- Запрещается работа двигателя на холостом ходу свыше 10 минут – вследствие низкого расходования горючего подшипник коленчатого вала охлаждается недостаточно, быстро перегревается, что может привести к заклиниванию.
- Для легкого запуска двигателя своевременно очищайте дренажное отверстие в крышке топливного бака, фильтрующий элемент.
- Из-за недостаточно прогрева двигателя, загрязненной свечи, неправильной установки наконечника провода высокого напряжения, двигатель может глохнуть или работать с перебоями.

 youtube.com/embed/jM1PDLrWHnM?feature=oembed»/>
youtube.com/embed/jM1PDLrWHnM?feature=oembed»/>Магнето – контроль системы зажигания
Тестирование системы производится визуально, с помощью щупа измеряют величину зазора между электродами. Для детального осмотра магнето снимают кожух и маховик, выполняют необходимые регулировки в соответствии с инструкцией.
При регулировке зазора, во избежание поломки, нельзя нажимать с силой на центральный электрод.
В инструкции по эксплуатации мотокультиватора Крот представлена очень подробная информация об устройстве агрегата, графике регламентных работ, настройке систем и механизмов, причинах неполадок и их устранении:
Видео обзор
Отзывы владельцев
Антон:
«Культиватор Крот у меня уже много лет выполняет все работы в хозяйстве. И не только по обработке земли, но и по перевозке урожая. Простой агрегат, проще наверное некуда, хорошие характеристики. Главное топливную смесь правильно готовить, расходные вовремя менять, приспособиться к работе. Если почва плотная и культиватор норовит выскочить наверх, углубляю сошник с усилием, а дальше сам идет нормально. Поломок серьезных за 7 лет не было, только профилактика».
Главное топливную смесь правильно готовить, расходные вовремя менять, приспособиться к работе. Если почва плотная и культиватор норовит выскочить наверх, углубляю сошник с усилием, а дальше сам идет нормально. Поломок серьезных за 7 лет не было, только профилактика».
Мотокультиватор Крот МК-1А
Благодаря высокой в своё время популярности мотокультиватор Крот МК-1А обзавёлся парой модификаций, которые несколько отличаются своими характеристиками, сохраняя всё же сходство с оригиналом. При этом сходство заключается не только в качестве работы, но и во внешних признаках. Встретить его можно где угодно, т.к. выпускается он с начала 80-х прошлого века.
Мотокультиватор Крот МК-1А в работе
Устройство мотокультиватора Крот МК-1А
По сравнению со своим предшественником мотокультиватор Крот МК-1А новой модификации имеет более эргономичную форму рукоятей, а также больший вес, что увеличивает проходимость при первичной обработке грунта. Помимо основного своего назначения – вспашки почвы, он со значительным успехом справляется с рядом других задач, среди которых числятся:
Помимо основного своего назначения – вспашки почвы, он со значительным успехом справляется с рядом других задач, среди которых числятся:
- Прополка сорняков;
- Окучивание грядок;
- Выкапывание картофеля;
- Сенокос;
- Перекачка воды;
- Перевозка небольших грузов.
Однако для выполнения данных операций потребуется приобретать дополнительно комплект навесного оборудования, состоящий из таких элементов:
- полольников;
- окучивателя;
- колёс с грунтозацепами;
- выкапывателя;
- плуга;
- косилки;
- насосной установки;
- тележки.
Такая оптимизация позволяет классифицировать инструмент не как мотокультиватор, а как мотоблок.
В зависимости от комплектации он может иметь 2-х или 4-хтактный двигатель. При использовании первого увеличивается моторесурс изделия, а во втором случае повышается силовая тяга, что гораздо удобнее на сложно проходимых участках. Мотор у мотокультиватора имеет воздушное охлаждение. Переключение передач у него – механическое, однако, в некоторых версиях движение может осуществляться только вперёд.
Переключение передач у него – механическое, однако, в некоторых версиях движение может осуществляться только вперёд.
Самый облегчённый вариант мотокультиватора Крот МК-1А имеет вес – 48 кг, что обеспечивает ему отличную мобильность в междурядьях.
Обзор мотокультиватора Крот МК-1А-02
Содержание
Мотокультиватор Крот МК-1А-02
Мотокультиватор “Крот” МК-1А-02 выпускается отечественным заводом АО «ММП им. В.В. Чернышева». Первые агрегаты были выпущены производителем в 80-х годах прошлого века. Эти устройства, хоть и отличались (характеристиками, устройством, принципом запуска и работы, прицепным оборудованием и т. д.) от современного моторизированного культиватора “Крот” МК-1А-02 , но были оценены по достоинству владельцами огородов, теплиц (хозяйств небольшого размера). С самого момента появления “Крот” стал показателем надежности, прочности, функциональности и высокой технологичности.
Описание
Современный агрегат с маркировкой МК-1А-02 был модернизирован, в результате чего увеличилась его мощность (2,6 л/с) и, соответственно, производительность. Двухтактный двигатель, работающий на смешанном топливе (бензин + масло) способен обеспечить бесперебойную работу мотокультиватора на протяжении долгого времени. Сравнительно небольшой вес (всего 48 кг) позволяет без особого труда транспортировать технику в любое место, требующее обработки мотокультиватором.
Двухтактный двигатель, работающий на смешанном топливе (бензин + масло) способен обеспечить бесперебойную работу мотокультиватора на протяжении долгого времени. Сравнительно небольшой вес (всего 48 кг) позволяет без особого труда транспортировать технику в любое место, требующее обработки мотокультиватором.
С помощью культиватора Крот можно производить следующие агротехнические работы на участке:
- вспашка грунта любой тяжести;
- прорезание почвы под посев;
- посадка картофеля;
- окучивание посаженных растений;
- прополка от сорняков;
- выкопка картофеля;
- покос;
- уборка снега;
- транспортировка грузов и т. д.
Все вышеописанные функции доступны со специальным навесным и прицепным оборудованием, которое можно купить или сделать самостоятельно ориентируясь на рисунки представленные в интернете.
Технические характеристики
| Параметр | Значение |
|---|---|
| Производитель | АО «ММП им. В.В. Чернышева» В.В. Чернышева» |
| Страна производитель | Россия |
| Диаметр фрез | 33 см. |
| Мощность | 2.6 л.с. |
| Количество скоростей | 1 вперед/0 назад |
| Реверс | Нет |
| Рулевая колонка | 1 положение |
| Ширина захвата | 35-60 см. |
| Глубина захвата | 25 см. |
| Вес | 48 кг. |
| Габариты | 1300x810x1060 мм. |
Инструкция по эксплуатации
Моторизированный культиватор МК-1А-02, кроме дополнительных функций, имеет основное предназначение — вспашка почвы. Для этого используются фрезы, которые насаживаются на вал, идущий от редуктора.
Настройка мотокультиватор перед вспашкой фрезой:
пневматические колеса приподнимаются, а движение агрегата происходит благодаря вращению фрез. В результате такого движения-рыхления происходит и вспахивание почвы.

Для почв более легких используется один-два комплекта фрез, для целинных грунтов — три (по 3 фрезы с каждой стороны мотокультиватора).
Существует и другой способ вспашки почвы — с помощью оборотного навесного плуга, который крепится на место сошника, вместо фрез устанавливаются металлические колеса.
В случае, когда необходимо провести другие агротехнические работы с агрегатом (прополка, посадка и т. д.), необходимо провести еще одно переоборудование мотокультиватора МК-1А-02.
Настройка мотокультиватора в зависимости от задач:
- При прополке растений с фрез снимаются ножи, а на их место устанавливаются полольники (эти приспособления имеют Г-образную форму). В случае с прополкой картофеля, установленный в задней части культиватора сошник будет выполнять роль окучника.
- Окучивание картофеля осуществляется без фрез, вместо которых устанавливаются металлические колеса, оснащенные грунтозацепами. Вместо сошника ставится окучник.
- Уборка картофеля производится следующим образом: впереди устанавливаются металлические колеса с грунтозацепами, а сзади цепляется навесное оборудование — картофелекопалка.
- Если вы желаете использовать культиватор МК-1А-02 в качестве газонокосилки — купите саму косилку и закрепите ее в передней части мотоагрегата. Для осуществления движения необходимо на валах редуктора закрепить пневмоколеса, а передачу необходимого крутящего момента обеспечат ремни, которые необходимо надеть на шкивы косилки и самого культиватора.
- Нужен насос — купите соответствующую насадку МНУ-2, закрепите ее на раме культиватора МК-1А-02 с помощью ременной передачи, не забыв отсоединить ремень тягового редуктора.
- Перевозка грузов до 200 кг возможна со специальной тележкой (прицепом), которая оснащена особым сцепным механизмом (поворотным). Транспортировка грузов осуществляется с помощью пневмоколес.

Краткий обзор навесного оборудования
Более детально ознакомится с навесным оборудованием для мотокультиватора Крот можно по этой ссылке.
Основные неисправности
Мотокультиватор МК-1А-02 “Крот” иногда требует ремонта. Рассмотрим основные поломки, которые требуют незамедлительного вмешательства оператора и быстрого ремонта агрегата:
- Отсутствие зажигания.
- Загрязнение воздушного фильтра.
- Поломка редуктора.
- Греется и глохнет мотор.
- Посторонние шумы в мотоблоке.
Причин поломки агрегата может быть несколько:
- Выход из строя или загрязнение свечи зажигания. В этом случае свечу необходимо либо хорошенько почистить и промыть, либо полностью заменить.
- Проблемы со шлангом подачи бензина.
- Нарушена изоляция высоковольтных проводов.
- Проблемы в соединении проводов (электросеть не контачит).
- Засорился фильтр. Его можно почистить, при сильных загрязнениях — заменить, благо, что эта запчасть всегда имеется в продаже.
- Пластины маховика клинят магнитопроводы магнето.
- Загрязнен карбюратор и потому машина глохнет (нагревается). Его необходимо тщательно вымыть, если неисправность не исчезает — заменить.
- При наличии посторонних шумов долейте масла.
- Заглох мотор — посмотрите, не закончилось ли топливо.
- Обратите внимание на редуктор, возможно следует подтянуть болты и гайки, или же заменить сальники.
 Его необходимо тщательно вымыть, если неисправность не исчезает — заменить.
Его необходимо тщательно вымыть, если неисправность не исчезает — заменить.Как вы заметили, чаще всего причиной быстрого нагревания двигателя на мотокультиваторе “Крот” МК-1А-02 является его загрязнение, а затем идут неполадки с электроникой и проводкой.
Содержите мотокультиватор МК-1А-02 “Крот” в чистоте и ремонт вам долго не понадобится.
Магнето МБ-1 схема для мотокультиватора “Крот”
Предлагаем вам ознакомиться с усовершенствованной схемой магнето к мотокультиватору МК-1 “Крот”.
Видео обзор
Чтобы увидеть мотокультиватор в работе, а также еще перед покупкой агрегата оценить его возможности, предлагаем посмотерть видео обзор.
Общий обзор мотокультиватора Крот
 youtube.com/embed/zUoPmNkZzrs?feature=oembed»/>
youtube.com/embed/zUoPmNkZzrs?feature=oembed»/>Отзывы владельцев
Что же говорят об этой агротехнике наши эксперты — обычные владельцы дачных участков и огородов, которым посчастливилось купить мотокультиватор “Крот”? Давайте ознакомимся с несколькими отзывами:
Иван Андреевич, 54 года
«В целом, мотокультиватор МК-1А-02 “Крот” мне очень нравится! Пользуюсь этой машиной уже 17 лет, агрегат зарекомендовал себя как надежная техника. За все время эксплуатации было несколько серьезных поломок: накрылся магнето — пришлось заменить, первое время глох движок, но я быстро усек в чем дело — тщательно прочистил карбюратор и впредь с неочищенным от пыли, грязи мотоблоком на работу не выходил. Пару раз приходилось в процессе вспашки поля подтягивать крепежные болты, машина сразу прекращала тарахтеть, как трещетка. Ну, что еще — сальники менял пару раз. Функционал впечатляющий, пришлось подкупить кое-какое оборудование, некоторое сделал сам (снегоуборщик). Мужики, рекомендую всем — не пожалеете!»
Василий, 39 лет
«Первый мотокультиватор Крот был еще у моего отца. Так что при покупке я уже наперед понимал, что покупаю. Меня все устраивает: и очень доступная цена, и хорошая производительность. Читал отзывы, что мол мотоблок для целины не годится — слабоват, отвечу — если пройтись несколькими этапами, да в разных направлениях — подчинится и целина! Купили с женой дом с большим участком, земля такая — думали ничего не получится! Но мотокультиватор приятно удивил, медленно, потихонечку взяли мы эту целину. Здесь главное знать как и не отступать!»
Мотокультиваторы Крот МК-1А
Культиватор Крот в модификации МК-1А-02 предназначен для предпосевной подготовки грунта, очистки огорода от сорняков, ухода за подросшими кустами и сбора урожая. Модель выделяется высокой маневренностью, которой удалось достичь благодаря небольшим габаритам и хорошей балансировке сельхозагрегата. Благодаря долговечному тяговитому двигателю мотокультиватор показывает внушительную выносливость, что позволяет использовать его для вспашки участков с тяжелой глинистой почвой.
В основу фирменной комплектации культиватора Крот вложен 2-тактный 1-цилиндровый силовой агрегат, укомплектованный системой воздушного охлаждения. Емкость рабочего цилиндра мотора составляет 60 см 3 , а номинальная скорость вращения коленвала – 5500–6500 об/мин.
За обогащение горючего воздухом и дальнейшую подачу готовой смеси в цилиндр двигателя в базовом устройстве мотокультиватора Крот отвечает карбюратор марки К-60В. В его устройство входит сухой воздухоочиститель, укомплектованный сменным фильтрующим элементом. Для заправки сельхозагрегата топливом в его устройстве предусмотрен бак емкостью 1,8 л. Двигатель модели работает от смеси, приготовленной из бензина АИ-80 или АИ-92, а также моторного масла марки М-8В1 в пропорции 20:1.
Для пуска встроенного мотора культиватор Крот МК-1А-02 укомплектован ручным несъемным стартером, штатная конструкция которого включает цельнометаллический вентилируемый барабан, усиленную возвратную пружину и трос с пластиковой рукояткой на его конце. Внутренние детали пускового механизма помещены в ударостойкий корпус с технологическими отверстиями для беспрепятственного охлаждения стартера воздухом.
Внутренние детали пускового механизма помещены в ударостойкий корпус с технологическими отверстиями для беспрепятственного охлаждения стартера воздухом.
Система зажигания мотокультиватора Крот состоит из бесконтактного электронного магнето марки МБ-1 и встроенной свечи зажигания А-17В. Сцепление модели представляет собой постоянно выключенный механизм, который приводится в действие при помощи мотоциклетной рукоятки.
В оснащение мотокультиватора Крот входит основной 2-ступенчатый редуктор, состоящий из пары шестерней и цепи. Для обеспечения стабильной работы узла его запчасти необходимо смазывать маслом марки ТАД-17.
Редуктор сельхозагрегата болтами крепится к раме, устройство которой включает 2 полурамы. На них закреплен кронштейн для возможности установки дополнительных приспособлений в задней части мотокультиватора, а также рукоятки для управления техникой.
Технические характеристики сельхозагрегатаЗаводские параметры модели Крот МК-1А-02 включают:
- мощность – 2,6 л. с.;
- количество скоростей – 1 передняя;
- ширина вспашки минимальная/максимальная – 35/60 см;
- глубина вспашки – до 25 см;
- вес в готовом к работе виде – 48 кг.
 с.;
с.;Несмотря на средние по современным меркам показатели мощности, мотокультиватор Крот показывает внушительную производительность. Так, во время фрезерования этот показатель достигает 150–200 с 2 /ч.
Инструкция по эксплуатации моделиСуществует ряд рекомендаций, придерживаясь которых пользователь мотокультиватора Крот сможет продлить сроки эксплуатации своего сельхозагрегата.
Производитель модели советует:
- использовать мотокультиватор только по назначению, не эксплуатировать его на целинных грунтах и крупных участках с каменистой почвой;
- заправлять культиватор качественным бензином и моторным маслом, смешивая компоненты только в пропорции 20:1;
- регулярно следить за количеством и состоянием смазок для деталей редуктора и трансмиссии, а также доливать жидкости, если их количество снижено;
- своевременно очищать воздушный и топливный фильтры, тщательно промывать карбюратор, трансмиссию и редуктор, а также продувать сжатым воздухом топливные трубки.

Во время эксплуатации мотокультиватора Крот МК-1А-02 нужно постоянно следить за состоянием его штатных фрез. Важно помнить, что работа с затупленными режущими органами приведет к повышению нагрузок на двигатель и снижению качества вспашки участка.
Мотокультиваторы Крот: обзор характеристик моделей
О мотокультиваторах марки Крот наслышаны многие фермеры и агрономы, ведь компания начала производство сельскохозяйственной техники еще в 80х годах и за весь период своей работы смогла доказать, что отечественные мотоблоки и мотокультиваторы ничем не уступают иностранной продукции по функциональности и надежности.
Мотокультваторы Крот – в чем причина популярности?
Эти культиваторы российского производства превосходят другие отечественные и некоторые импортные аналоги практически по всем параметрам. Такая техника позволяет установить плуг, фрезы и различные окучники. Эти модели успешно справляются с обработкой целины и тяжелых глинистых грунтов.
Как показывает практика, лучше всего эти агрегаты подходят для работы на участках, площадью не более 10 соток.
Отличительные особенности:
- Наличие мощных двухтактных моторов, преимущественно Honda, мощностью от 2 до 8 л. с.;
- Комплектация качественными редукторами, выдерживающими высокие нагрузки;
- Наличие эргономичных ручек и удачное расположение органов управления;
- Компактные габариты и небольшой вес;
- Качественная заточка фрез;
- Глубина обработки 0 от 12 до 25 см;
- Ширина охватываемой полосы – 30–110 см.
Самой компактной моделью считается агрегат «Крот М», обладающий мощностью в 5 л. с. Он подходит для вспашки небольших участков с применением широкого спектра навесного оборудования. Если обрабатываемый участок имеет большую площадь, то стоит рассмотреть представителей ряда МК – они более мощные и надежные.
Мотокультиваторы Крот: детально рассматриваем самые популярные модели
Сегодня компания представляет на рынке невероятное разнообразие вариантов, поэтому, чтобы оценить все положительные особенности продукции, следует познакомиться с несколькими представителями бренда.
Культиватор крот МК-9-02
Этот агрегат считается одним из самых современных, поскольку он оснащен 4х-тактным двигателем Hammermann с верхним расположением кулачков. Немалая мощность и продуманность всех деталей превращают его в очень достойного соперника для приспособлений иностранного производства.
Технические характеристики:
- Мощность двигателя – 5, 5 л. с.;
- Вместительность топливного бака – 3, 6 л.;
- Запуск – ручной;
- Система зажигания – транзисторное магнето;
- Используемое топливо – автомобильный бензин АИ-92, АИ-95;
- Необходимое масло – SF, SG, SAE 30, SAE 10W-40;
- Вес – 53, 2 кг.
Благодаря воздушной системе охлаждения удается продлить срок эксплуатации культиватора, тем самым избежав перегреваний и порчи деталей из-за высокой температуры. В том случае, если появляется необходимость в использовании ходоуменьшителей, можно пускать в ход колеса большого диаметра, которые будут оборачиваться гораздо медленней комплектационных и способствовать замедлению агрегата.
Мотокультиватор Крот МК-5-01
Благодаря удивительному сочетанию отточенной годами конструкционной специфики компании Крот и надежности японского двигателя Хонда, удалось создать устройство, отображающее самые выгодные особенности обеих организаций.
Технические характеристики:
- Мощность двигателя – 4 л. с.;
- Количество передач – 2;
- Вместительность топливного бака – 1,7 л;
- Результативность – 150-200 м 2 /час;
- Используемое топливо – автомобильный бензин АИ-92, АИ-95;
- Масса – 48 кг.
Повысить уровень производительности и расширить и без того широкий спектр реализуемых задач поможет специальное навесное оборудование, которое можно приобрести в магазинах или же попытаться сделать самостоятельно, ведь нередко простые самоделки оказываются более продуктивными чем дорогие вспомогательные детали.
Культиватор Крот МК-4-01
Агрегат дополнен двигателем известной американской компании «Briggs & Stratton», поэтому можно с уверенностью сказать, что мощности детали хватит на выполнение многих задач. 3,6 лошадиных сил будет достаточно для обработки небольшого земельного участка или внушительной приусадебной территории, рассчитанной на высадку овощей и корнеплодов.
3,6 лошадиных сил будет достаточно для обработки небольшого земельного участка или внушительной приусадебной территории, рассчитанной на высадку овощей и корнеплодов.
Технические характеристики Крот МК-4-01:
- Вместительность топливного бака – 1,8 л;
- Тип топлива – бензин АИ-93, АИ-92;
- Производительность в процессе фрезерования – 150-200 м 2 /час.
- Глубина обработки – до 250 мм.;
- Вес – 50 кг.
Немало радует, что такие агрегаты сопровождаются специальными инструкциями, где представлена информация по ремонту и подбору запчастей. Более того, подобные руководства подскажут, как измерить диаметр вала, заменить его, «привести в чувства» культиватор и все, что связано с ним.
Мотокультиватор Крот 1А
Мотокультиватор Крот МК 1А относится к наиболее популярным представителям сельхозтехники. Из-за высокого спроса на агрегат, производитель расширил модельный ряд этого семейства. Сегодня техника встречается в нескольких комплектациях, что дает возможность каждому из покупателей выбрать наиболее подходящий для себя вариант.
Современный мотокультиватор Крот 1А снабжается рукояткой с эргономичной формой. Модель весит немного больше своего предшественника, что повышает ее проходимость в процессе первичной обработки почвы. Помимо вспашки грунта, агрегат успешно справляется со многими другими задачами. Так, модель используется для:
- Прополки грядок и удаления сорняков;
- Окучивания грядок;
- Сбора картофеля;
- Покоса сена;
- Перекачки воды;
- Транспортировки небольших грузов.
Конечно, для всех этих работ необходимо отдельно приобретать нужный инвентарь. Окучник, колеса с грунтозацепами, картофелекопалка, плуги, косилка, тележка.
В зависимости от своей комплектации, модель может снабжаться 2-или 4-тактным мотором. Эксплуатация первого увеличивает ресурс агрегата, а использование второго повышает силовую тягу – это более удобно на заросших участках. Двигатель культиватора работает под воздушным охлаждением. Трансмиссия механическая, некоторые агрегаты могут двигаться только вперед. Самая легкая комплектация имеет вес в 48 кг. Благодаря этому агрегат отличается маневренностью на узких участках.
Самая легкая комплектация имеет вес в 48 кг. Благодаря этому агрегат отличается маневренностью на узких участках.
Культиватор Крот 1А обладает улучшенной конструкцией, что положительно влияет на его технические показатели. Агрегат отличается повышенной надежностью запчастей, отличным качеством сборки и наличием защиты от загрязнений и попадания воды.
Технические характеристики культиватора Крот 1А:
- Крутящий момент двигателя составляет 5000–6500 об./мин.;
- Мощность мотора – 2,6 л. с.;
- Ширина обработки грунта регулируется, и может составлять от 35 до 60 см.;
- Одна передняя скорость;
- Глубина вспашки – не более 25 см.;
- Средняя производительность составляет 170–200 квадратных метров за час работы.
Одним из преимуществ модели является ее небольшие габариты. Высота агрегата составляет 130, ширина – 81, а высота – 106 см. Благодаря сравнительно скромным размерам, агрегат успешно используется для работки грунта на клумбах, в парниках и небольших участках с обильными посадками.
Мотокультиватор Крот МК 1А 02
Рабочие показатели этой комплектации практически полностью идентичны с предыдущей моделью. Тем не менее, этот агрегат имеет некоторые конструктивные отличия. К ним относится:
- Доработанная система сцепления – благодаря ей владельцу легче запускать агрегат. Кроме того, во время пуска мотор потребляет на 20 % меньше топлива;
- Улучшенная трансмиссия – модель работает только на одной передней скорости, однако передача оборотов на вал происходит по укороченному маршруту, благодаря чему модель передвигается более уверенно;
- Стабилизированная система охлаждения – мотор модели будет меньше перегреваться даже при очень сильных нагрузках. Таким образом, двигатель тратит меньше времени на охлаждение, а его обслуживание нужно выполнять реже;
- Аппарат снабжается защитными пластинами, которые не дают воде и грязи попадать внутрь мотора и редуктора.
Спектр работ у модели МК 1А 02 достаточно широкий. Агрегат успешно используется для вспашки редко обрабатываемой почвы, он легко справляется с высокими сорняками, способен перекачивать большие объемы воды и косить сено.
Возникли неисправности с культиватором Крот? Что делать и как быть?
Представленная на нашем сайте универсальная инструкция по эксплуатации подойдет для решения проблем, связанных со всеми моделями производителя Крот. Отыскать достойные альтернативы оригинальным запчастям не сложно, ведь нередко в магазинах имеются необходимые детали. Стоит отметить, что к мотокультиваторам Крот зачастую подходят и китайские элементы.
Проблема
Стоит всегда помнить, что руководство поможет разобраться с возникшей ситуации и позволит избавиться от проблемы даже своими руками.
Расход топлива культиваторами Крот
Большинство современных моделей оснащаются центробежным регулятором частоты вращения коленвала, который подключается непосредственно к мотору. Это позволяет снизить расход топлива моделями Крот. На понижение расхода также влияет воздушный фильтр, улучшенный карбюратор и наличие реверса, дающего возможность двигаться в заднем направлении.
В большинстве моделей расход топлива не превышает 1 л. смеси за час работы. Почти все агрегаты работают на низкооктановом бензине марки АИ-76. Бензин следует перемешивать с маслом М-12 ТП или МГ-8А. Объем топливного бака составляет 1,8 л.
Правила эксплуатации культиваторов Крот
В процессе использования техники производитель советует придерживаться определенных правил. К ним относиться:
К ним относиться:
- Двигатель должен разогреваться не менее 5 минут;
- Если агрегат не запускается с пятой подряд попытки, первым делом нужно проверить целостность троса сцепления;
- Использовать для заправки только свежую топливную смесь;
- Не держать культиватор без работы больше 3 месяцев;
- Своевременно менять сальники и смазывать редуктор.
- 1 раз в три месяца чистить или менять воздушный фильтр.
Все эти советы помогут продлить жизнь технике. Если вы заметите какую-либо неисправность, можно попытаться устранить ее самостоятельно. Однако если опыта в ремонте культиватора нет, то лучше не рисковать, и отвезти агрегат прямо к мастеру.
1 Отзыв
Отечественные бензиновые культиваторы Крот являются достаточно известными, так как они используются многими владельцами участков на протяжении 30 лет. Конструкция таких изделий дает возможность использовать различное навесное оборудование для выполнения работ по обработке земли.
Мотокультиватор крот мк 1а 02 в Архангельске: 16-товаров: бесплатная доставка, скидка-23% [перейти]
Партнерская программаПомощь
Архангельск
Каталог
Каталог Товаров
Одежда и обувь
Одежда и обувь
Стройматериалы
Стройматериалы
Текстиль и кожа
Текстиль и кожа
Здоровье и красота
Здоровье и красота
Продукты и напитки
Продукты и напитки
Детские товары
Детские товары
Электротехника
Электротехника
Дом и сад
Дом и сад
Промышленность
Промышленность
Мебель и интерьер
Мебель и интерьер
Сельское хозяйство
Сельское хозяйство
Все категории
ВходИзбранное
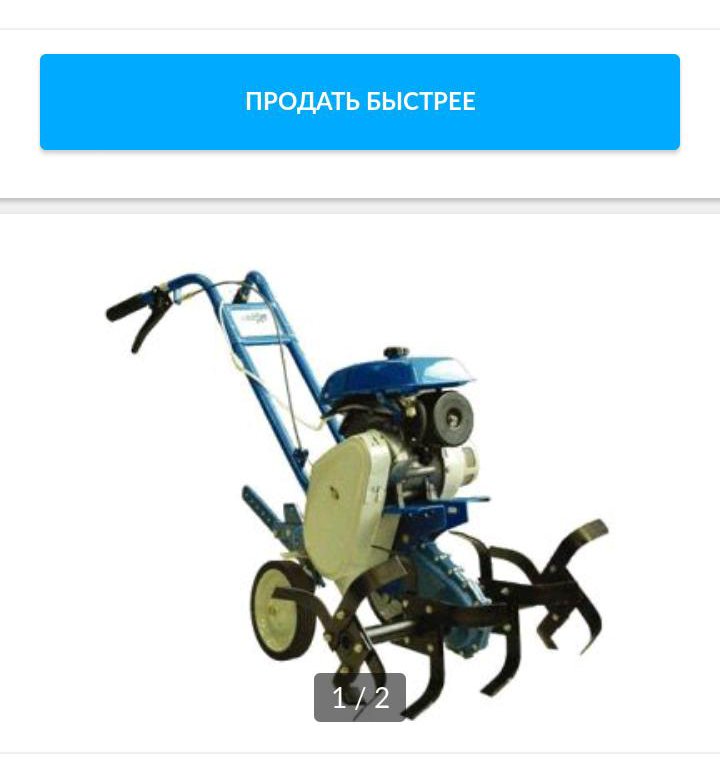
Мотокультиватор крот мк 1а 02
40 350
Мотокультиватор «Нева» МК-100Р-ZS, GB210, 6 фрез, реверс Тип: культиватор, Бренд: Нева, Тип
ПОДРОБНЕЕЕще цены и похожие товары
 regmarkets.ru/listpreview/idata2/c8/9f/c89f0f589b0e8eb03a55664e2bc0bd1e.jpg»>
regmarkets.ru/listpreview/idata2/c8/9f/c89f0f589b0e8eb03a55664e2bc0bd1e.jpg»>1 776
2079
Стартер для мотокультиватора КРОТ (SDDh257) Тип: Стартер для бензоинструмента, Размер: Длина 25.000
В МАГАЗИНЕще цены и похожие товары
305
305
Трос сцепления для мотокультиватора Крот Тип: Ремкомплект для бензоинструмента, Размер: Длина
В МАГАЗИНЕще цены и похожие товары
390
420
Прокладки мотокультиватор Крот набор Тип: Культиватор ручной, Размер: Длина 15.000 Ширина 11.000
В МАГАЗИНЕще цены и похожие товары
Культиваторы ( пара ) к мотокультиватору Крот 2000000023267
В МАГАЗИНЕще цены и похожие товары
Ручной стартер для мотокультиватора Крот A71010 101046
В МАГАЗИНЕще цены и похожие товары
Культиваторы Целина ( пара ) 5-ми зубые к мотокультиваторам ( d=30 ) Крот , Тарпан
ПОДРОБНЕЕЕще цены и похожие товары
Культиваторы (пара) к м/к Крот (Пермь) CARVER 01. 015.00014/4
015.00014/4
ПОДРОБНЕЕЕще цены и похожие товары
Катушка зажигания для мотокультиватора крот Тип: катушка зажигания, Производитель: Без бренда, Вес:
ПОДРОБНЕЕЕще цены и похожие товары
49 990
Мотоблок бензиновый BR-135GAPRO с пониженной передачей / мотокультиватор / культиватор для вспашки земли / брайт / сельскохозяйственная машина
ПОДРОБНЕЕЕще цены и похожие товары
Ремень клиновой ширина-13мм для мотоблока Крот Тип ремня: клиновый, Ширина ремня: 13мм
ПОДРОБНЕЕЕще цены и похожие товары
 jpg»>
jpg»>306
420
A-710 Ремень клиновой.Для мотокультиватока «Крот«, «Сибиряк» Производитель: Без бренда, Тип ремня:
ПОДРОБНЕЕЕще цены и похожие товары
Культиваторы (пара) к м/к Крот (Пермь)
ПОДРОБНЕЕЕще цены и похожие товары
Шкив редуктора большой, металлический, 2-х ручейковый мотокультиватора Крот Тип: шкив,
ПОДРОБНЕЕЕще цены и похожие товары
Шкив редуктора, большой, металлический, штампованный, 1-ручейковый мотокультиватора Крот Тип: шкив,
ПОДРОБНЕЕЕще цены и похожие товары
Редуктор в сборе для мотокультиватора (без крепления руля) Тип: редуктор (в сборе), Производитель:
ПОДРОБНЕЕЕще цены и похожие товары
 regmarkets.ru/listpreview/images3/d1/3e/d13e8c6d6f5f712e81b0abb35ef5a917.jpg»>
regmarkets.ru/listpreview/images3/d1/3e/d13e8c6d6f5f712e81b0abb35ef5a917.jpg»>Ремень клиновой A-750 для культиваторов «Крот» Тип: ремень, Назначение: для культиватора, Длина:
ПОДРОБНЕЕЕще цены и похожие товары
Ремень привода А-900 для культиватора Крот
ПОДРОБНЕЕЕще цены и похожие товары
Пропольники с дисками защиты на мотокультиваторы типа Крот, Тарпан. На диаметр вала — 25 мм.
ПОДРОБНЕЕЕще цены и похожие товары
 regmarkets.ru/listpreview/images3/91/f1/91f141e1d6bc1f65da0c78bbbd4bb171.jpg»>
regmarkets.ru/listpreview/images3/91/f1/91f141e1d6bc1f65da0c78bbbd4bb171.jpg»>Зажигание электронное МБ-1 на мотокультиватор Крот (Совек) Тип: культиватор
ПОДРОБНЕЕЕще цены и похожие товары
37 900
Культиватор нева МК100Р-ZS GB210 Тип: культиватор, Бренд: Нева, Тип двигателя: бензиновый
ПОДРОБНЕЕЕще цены и похожие товары
Ремень клиновой А-710 Lp / 680 Li — (для культиватора Крот) (5 шт.) Тип: ремень, Производитель: Без
ПОДРОБНЕЕЕще цены и похожие товары
 jpg»>
jpg»>15 700
Двигатель Briggs & Stratton 550 Series (4л.с.) для Крота Тип: бензиновый, Производитель: BRIGGS &
ПОДРОБНЕЕЕще цены и похожие товары
Ремень (13Х925) для мотокультиватора «Крот«
ПОДРОБНЕЕЕще цены и похожие товары
Редуктор в сборе для мотокультиватора (без крепления руля) Тип: редуктор (в сборе), Производитель:
ПОДРОБНЕЕЕще цены и похожие товары
Кожух защитный ремня мотокультиватора Крот Производитель: Не определен, Назначение: для культиватора
ПОДРОБНЕЕЕще цены и похожие товары
Плуг к мотокультиваторам Крот б/сцепки (Пермь) стойка 10*500 мм , ширина отвала 190 мм Тип: плуг
ПОДРОБНЕЕЕще цены и похожие товары
 regmarkets.ru/listpreview/images3/72/40/724031ce9bfe199c442d6f18e0ca809e.jpg»>
regmarkets.ru/listpreview/images3/72/40/724031ce9bfe199c442d6f18e0ca809e.jpg»>Ремень клиновой A-750 для мотоблоков и культиваторов Крот
ПОДРОБНЕЕЕще цены и похожие товары
Окучник роторный (активный) для мотокультиваторов с диаметром вала 25 мм. Сделан в России Тип:
ПОДРОБНЕЕЕще цены и похожие товары
Комплект расходников «Макси» для мотокультиваторов Крот
ПОДРОБНЕЕЕще цены и похожие товары
Грунтозацепы (пара) мотокультиватора Крот (Dнар. 290мм) Тип: грунтозацеп
290мм) Тип: грунтозацеп
ПОДРОБНЕЕЕще цены и похожие товары
Ремень А750 для культиватора Крот
ПОДРОБНЕЕЕще цены и похожие товары
Стартер мотокультиватора Крот в сборе
ПОДРОБНЕЕЕще цены и похожие товары
Культиваторы (пара) к м/к Крот разборные «Стандарт» CARVER 01.015.00014/5
ПОДРОБНЕЕЕще цены и похожие товары
16 650
Культиватор бензиновый КАЛИБР МК-4,5, 4.49 л.с. Тип: культиватор, Бренд: КАЛИБР, Тип двигателя:
ПОДРОБНЕЕЕще цены и похожие товары
Ручной стартер 101046 для мотокультиватора Крот A71010 Тип: стартер
ПОДРОБНЕЕЕще цены и похожие товары
Цилиндр мотокультиватора Крот Производитель: Без бренда
ПОДРОБНЕЕЕще цены и похожие товары
 regmarkets.ru/listpreview/images3/ce/6a/ce6a6d0a0ffbe4a5639cb6073e23e86a.jpg»>
regmarkets.ru/listpreview/images3/ce/6a/ce6a6d0a0ffbe4a5639cb6073e23e86a.jpg»>A-900 Ремень клиновой. Для мотокультиватора «Крот» Тип: ремень, Производитель: Без бренда, Вес:
ПОДРОБНЕЕЕще цены и похожие товары
Плуг для мотокультиватора крот, ЗУБР, FORZA (универсальный) Тип: плуг
ПОДРОБНЕЕЕще цены и похожие товары
 regmarkets.ru/listpreview/images3/3c/73/3c73c3456eab380257e4ffca97b0d2a4.jpg»>
regmarkets.ru/listpreview/images3/3c/73/3c73c3456eab380257e4ffca97b0d2a4.jpg»>Стартер для мотокультиватора крот
ПОДРОБНЕЕЕще цены и похожие товары
Удлинители грунтозацепов для мотокультиваторов Крот (комплект) Тип: удлинитель
ПОДРОБНЕЕЕще цены и похожие товары
57 500
KROTOF TLR1060L Мотокультиватор бензиновый с электрозапуском (13 л.с., передачи 3 вперед / 1 назад)
ПОДРОБНЕЕЕще цены и похожие товары
23 651
Мотокультиватор Sterwins Model-1 3,5 л/с Тип: культиватор
ПОДРОБНЕЕЕще цены и похожие товары
Шкив ведущий, 2-х ручейковый мотокультиватора Крот Тип: шкив, Производитель: Без бренда, Ширина:
ПОДРОБНЕЕЕще цены и похожие товары
Шкив для мотокультиватора крот диаметр 19мм Производитель: Kimotozip
ПОДРОБНЕЕЕще цены и похожие товары
Прокладки мотокультиватор Крот набор Тип: культиватор
ПОДРОБНЕЕЕще цены и похожие товары
 regmarkets.ru/listpreview/images3/8d/0d/8d0d97cb508c69bbb4b98bc5938146b3.jpg»>
regmarkets.ru/listpreview/images3/8d/0d/8d0d97cb508c69bbb4b98bc5938146b3.jpg»>Колеса транспортировочные для мотокультиватора типа Крот, Тарпан. Сделаны на Кубани. Тип: колеса
ПОДРОБНЕЕЕще цены и похожие товары
Комплект расходников «Мини» для мотокультиваторов Крот
ПОДРОБНЕЕЕще цены и похожие товары
Плуг для культиватора «Тарпан» Тип: плуг, Производитель: ТАРПАН
ПОДРОБНЕЕЕще цены и похожие товары
Культиваторы ( пара ) к мотокультиватору Крот 2000000023267
ПОДРОБНЕЕЕще цены и похожие товары
 jpg»>
jpg»>Ремень клиновой (2шт.) A-750 для мотоблоков и культиваторов Крот
ПОДРОБНЕЕЕще цены и похожие товары
Ремень Крот А750 Россия
ПОДРОБНЕЕЕще цены и похожие товары
Ремень клиновой A-750 для мотоблоков и культиваторов Крот
ПОДРОБНЕЕЕще цены и похожие товары
2 страница из 5
Мотокультиватор крот мк 1а 02 в Пятигорске: 21-товар: бесплатная доставка, скидка-18% [перейти]
Партнерская программаПомощь
Пятигорск
Каталог
Каталог Товаров
Одежда и обувь
Одежда и обувь
Стройматериалы
Стройматериалы
Текстиль и кожа
Текстиль и кожа
Здоровье и красота
Здоровье и красота
Детские товары
Детские товары
Продукты и напитки
Продукты и напитки
Электротехника
Электротехника
Дом и сад
Дом и сад
Сельское хозяйство
Сельское хозяйство
Мебель и интерьер
Мебель и интерьер
Торговля и склад
Торговля и склад
Все категории
ВходИзбранное
Мотокультиватор крот мк 1а 02
40 350
Мотокультиватор «Нева» МК-100Р-ZS, GB210, 6 фрез, реверс Тип: культиватор, Бренд: Нева, Тип
ПОДРОБНЕЕЕще цены и похожие товары
Ручной стартер для мотокультиватора Крот A71010 101046
В МАГАЗИНЕще цены и похожие товары
 regmarkets.ru/listpreview/idata2/7e/f0/7ef0e3b4d9cfde9045d3c44cfe02ac3d.jpg»>
regmarkets.ru/listpreview/idata2/7e/f0/7ef0e3b4d9cfde9045d3c44cfe02ac3d.jpg»>305
305
Трос сцепления для мотокультиватора Крот Тип: Ремкомплект для бензоинструмента, Размер: Длина
В МАГАЗИНЕще цены и похожие товары
1 776
2079
Стартер для мотокультиватора КРОТ (SDDh257) Тип: Стартер для бензоинструмента, Размер: Длина 25.000
В МАГАЗИНЕще цены и похожие товары
Культиваторы ( пара ) к мотокультиватору Крот 2000000023267
В МАГАЗИНЕще цены и похожие товары
390
420
Прокладки мотокультиватор Крот набор Тип: Культиватор ручной, Размер: Длина 15. 000 Ширина 11.000
000 Ширина 11.000
В МАГАЗИНЕще цены и похожие товары
Шестерня большая поз. A 28 для культиватора Patriot Крот II (004517009) PATRIOT запчасти
ПОДРОБНЕЕЕще цены и похожие товары
1 610
1850
Ручной стартер для мотокультиватора Крот A71010 101046
ПОДРОБНЕЕЕще цены и похожие товары
Ремень клиновой A-750 для культиваторов «Крот» Тип: ремень, Назначение: для культиватора, Длина:
ПОДРОБНЕЕЕще цены и похожие товары
Ручной стартер 101046 для мотокультиватора Крот A71010 Тип: стартер
ПОДРОБНЕЕЕще цены и похожие товары
Плуг для культиватора «Тарпан» Тип: плуг, Производитель: ТАРПАН
ПОДРОБНЕЕЕще цены и похожие товары
 regmarkets.ru/listpreview/images3/8f/8a/8f8a6a1dbd154f3aedb5c2186d0d0fd4.jpg»>
regmarkets.ru/listpreview/images3/8f/8a/8f8a6a1dbd154f3aedb5c2186d0d0fd4.jpg»>A-750 Ремень клиновой. Для мотокультиватора «Крот» Тип: ремень, Производитель: Без бренда, Вес:
ПОДРОБНЕЕЕще цены и похожие товары
306
420
A-710 Ремень клиновой.Для мотокультиватока «Крот«, «Сибиряк» Производитель: Без бренда, Тип ремня:
Ручной стартер подходит для мотокультиватора Крот
ПОДРОБНЕЕЕще цены и похожие товары
49 990
Мотоблок бензиновый BR-135GAPRO с пониженной передачей / мотокультиватор / культиватор для вспашки земли / брайт / сельскохозяйственная машина
ПОДРОБНЕЕЕще цены и похожие товары
 regmarkets.ru/listpreview/images3/9b/6b/9b6bc43f6b568dd1330386ced32bc858.jpg»>
regmarkets.ru/listpreview/images3/9b/6b/9b6bc43f6b568dd1330386ced32bc858.jpg»>Ремень клиновой ширина-13мм для мотоблока Крот Тип ремня: клиновый, Ширина ремня: 13мм
ПОДРОБНЕЕЕще цены и похожие товары
Ролик натяжной широкий для мотокультиватора Крот
ПОДРОБНЕЕЕще цены и похожие товары
A-900 Ремень клиновой. Для мотокультиватора «Крот» Тип: ремень, Производитель: Без бренда, Вес:
ПОДРОБНЕЕЕще цены и похожие товары
Комплект навесного оборудования DDE для «Крот» (02. 05.10-12.02.20-00.0425) Тип: комплект навесного
05.10-12.02.20-00.0425) Тип: комплект навесного
ПОДРОБНЕЕЕще цены и похожие товары
Культиваторы (пара) к м/к
ПОДРОБНЕЕЕще цены и похожие товары
Культиваторы (пара) к м/к Крот (Пермь) CARVER 01.015.00014/4
ПОДРОБНЕЕЕще цены и похожие товары
Культиваторы (пара) к м/к Крот (Пермь)
ПОДРОБНЕЕЕще цены и похожие товары
Ремень 13х925 зубчатый культиватор Крот Производитель: AEZ
ПОДРОБНЕЕЕще цены и похожие товары
Ремень (13Х925) для мотокультиватора «Крот» Страна-производитель: Китай
ПОДРОБНЕЕЕще цены и похожие товары
Ручной стартер мотокультиватора МК—1А Крот Производитель: Без бренда
ПОДРОБНЕЕЕще цены и похожие товары
Кожух защитный ремня мотокультиватора Крот Производитель: Не определен, Назначение: для культиватора
ПОДРОБНЕЕЕще цены и похожие товары
 regmarkets.ru/listpreview/images3/60/5c/605cc1d8a8d0262330d6c864aa26d1ae.jpg»>
regmarkets.ru/listpreview/images3/60/5c/605cc1d8a8d0262330d6c864aa26d1ae.jpg»>Ремень клиновой A-750 для мотоблоков и культиваторов Крот
ПОДРОБНЕЕЕще цены и похожие товары
Ремень А750 для культиватора Крот
ПОДРОБНЕЕЕще цены и похожие товары
Редуктор в сборе для мотокультиватора (без крепления руля) Тип: редуктор (в сборе), Производитель:
ПОДРОБНЕЕЕще цены и похожие товары
Цепь редуктора ПР15,875-2300-2 (42 звена) мотокультиватора Крот Тип: редуктор, Производитель: Без
ПОДРОБНЕЕЕще цены и похожие товары
 regmarkets.ru/listpreview/images3/de/7f/de7fc0a2b8bd549e8ba2da773423e7a8.jpg»>
regmarkets.ru/listpreview/images3/de/7f/de7fc0a2b8bd549e8ba2da773423e7a8.jpg»>Ремень клиновой (2шт.) A-710 для мотоблоков и культиваторов Крот, Сибиряк
ПОДРОБНЕЕЕще цены и похожие товары
Шкив для мотокультиватора крот диаметр 19мм
ПОДРОБНЕЕЕще цены и похожие товары
Маховик для культиватора Крот 150106900
ПОДРОБНЕЕЕще цены и похожие товары
Ремень Крот А750 ММ
ПОДРОБНЕЕЕще цены и похожие товары
 jpg»>
jpg»>Стартер для мотокультиватора крот
ПОДРОБНЕЕЕще цены и похожие товары
Окучник мотокультиватора Крот без сцепки Тип: окучник
Окучник роторный (активный) для мотокультиваторов с диаметром вала 25 мм. Сделан в России Тип:
ПОДРОБНЕЕЕще цены и похожие товары
Катушка зажигания для мотокультиватора крот Тип: катушка зажигания, Производитель: Без бренда, Вес:
ПОДРОБНЕЕЕще цены и похожие товары
Ремень клиновой А-710 Lp / 680 Li — (для культиватора Крот) Тип: ремень, Производитель: Без бренда,
ПОДРОБНЕЕЕще цены и похожие товары
 regmarkets.ru/listpreview/images3/90/4f/904ff9d80af66dcf165f83e290dc7af5.jpg»>
regmarkets.ru/listpreview/images3/90/4f/904ff9d80af66dcf165f83e290dc7af5.jpg»>27 500
Кротов TLR1150 Мотокультиватор Тип: культиватор, Бренд: Krotof, Мощность Вт: 5150.84621Вт
ПОДРОБНЕЕЕще цены и похожие товары
Редуктор для мотокультиватора МК Крот (без реверса) (15003000) Тип: редуктор
ПОДРОБНЕЕЕще цены и похожие товары
Ремень клиновой A-750 для мотоблоков и культиваторов Крот
ПОДРОБНЕЕЕще цены и похожие товары
35 500
Культиватор бензиновый Нева МК100Р-Kasei (168F), 5 л. с. Тип: культиватор, Бренд: Нева, Тип
с. Тип: культиватор, Бренд: Нева, Тип
ПОДРОБНЕЕЕще цены и похожие товары
Шкив редуктора большой, металлический, 2-х ручейковый мотокультиватора Крот Тип: шкив,
ПОДРОБНЕЕЕще цены и похожие товары
Ремень клиновой А-710 Lp / 680 Li — (для культиватора Крот) (5 шт.) Тип: ремень, Производитель: Без
ПОДРОБНЕЕЕще цены и похожие товары
Ремень клиновой А-710 Lp / 680 Li — (для культиватора Крот) (2 шт.) Тип: ремень, Производитель: Без
ПОДРОБНЕЕЕще цены и похожие товары
Плуг для мотокультиватора крот, ЗУБР, FORZA (универсальный) Тип: плуг
ПОДРОБНЕЕЕще цены и похожие товары
Стартер мотокультиватора Крот (Китай)
ПОДРОБНЕЕЕще цены и похожие товары
Культиваторы ( пара ) к мотокультиватору Крот 2000000023267
ПОДРОБНЕЕЕще цены и похожие товары
28 900
Культиватор бензиновый Нева МК-70-Kasei, 5 л. с. Тип: культиватор, Бренд: Нева, Тип двигателя:
с. Тип: культиватор, Бренд: Нева, Тип двигателя:
ПОДРОБНЕЕЕще цены и похожие товары
Шкив редуктора, большой, металлический, штампованный, 1-ручейковый мотокультиватора Крот Тип: шкив,
ПОДРОБНЕЕЕще цены и похожие товары
Ремень клиновой A-750 для мотоблоков и культиваторов Крот
ПОДРОБНЕЕЕще цены и похожие товары
Культиваторы Целина ( пара ) 5-ми зубые к мотокультиваторам ( d=30 ) Крот , Тарпан
ПОДРОБНЕЕЕще цены и похожие товары
2 страница из 5
Мотоблок, культиватор Крот, модели — технические характеристики
Культиватор Крот появился на рынке сельхозтехники ещё в прошлом веке. Агрегат подходит для дачных участков и небольших фермерских хозяйств. Постепенно техника подверглась модернизации, увеличивалась её функциональность благодаря большей мощности и возможности использовать навесное оборудование.
Агрегат подходит для дачных участков и небольших фермерских хозяйств. Постепенно техника подверглась модернизации, увеличивалась её функциональность благодаря большей мощности и возможности использовать навесное оборудование.
Содержание
- 1 Преимущества культиваторов Крот
- 2 Мотокультиватор Крот: устройство агрегата
- 3 Мотокультиватор Крот МК 1А 01
- 4 Мотокультиватор Крот МК 1А 02
Преимущества культиваторов Крот
Благодаря постоянной модификации и совершенствованию мотоблоков Крот можно осуществлять многие работы по обработке грунта. Вне зависимости от модификации техника остаётся доступной для покупки.
Преимущества использования агрегатов Крот:
- Надёжность;
- Простота при эксплуатации;
- Наличие удобных рукояток, позволяющих долго работать без чувства усталости;
- Мотокультиваторы комплектуются фрезами;
- Применение различного навесного оборудования;
- Доступная цена.

К недостатку культиваторов Крот относится небольшая ширина обработки полосы.
Мотокультиватор Крот: устройство агрегата
Большинство первых моделей с двухтактным одноцилиндровым двигателем. Современные мотоблоки Крот уже с четырёхтактными импортными двигателями. Они обладают достаточной мощностью для решения разных задач.
К комплектации мотокультиватора Крот должны быть следующие элементы:
- Ручка сцепления;
- Ручка настройки дросселя;
- Управление заслонкой карбюратора.
Техническая комплектация мотоблока:
- Мотор от 2,6 л. с. и объёмом от 60 см2;
- Электрозажигание бесконтактного типа;
- Стартер для ручного старта;
- Карбюратор «К-60В»;
- Топливный бак с вместительностью от 1,8 литра;
- Воздушные фильтры для очистки воздуха.
Оптимально использовать автол М8В, как смазку для двигателя и одноступенчатого редуктора.
На некоторых устройствах предусмотрено наличие задней передачи для удобного перемещения по участку и эффективного вспахивания грунтов. Навесы крепятся на входных валах редуктора.
Навесы крепятся на входных валах редуктора.
При транспортировке культиватора Крот на него устанавливаются колеса, которые удобно снимать перед тем, как приступить к работам.
Среди всей линейки продукции самыми актуальными остаются агрегаты Крот МК-1А 01 и 02, с характеристиками которых стоит ознакомиться детальнее.
Мотокультиватор Крот МК 1А 01
Крот МК-1А 01 с продуманной рукояткой для упрощения управления агрегатом и повышенной массой, которая обеспечивает отличную манёвренность при обработке целины.
Кроме ключевых работ по вспашке грунта мотокультиватор справиться со многими задачами:
- Покос/транспортировка сена;
- Транспортировка грузов небольшого веса;
- Окучивание грядок;
- Выкапывание клубневых культур;
- Уборка сорняков;
- Перекачивание воды.

Технические характеристики данной модели:
- Мотор с мощностью 2,6 л. с.;
- Топливный бак – 1,8 литра;
- Наличие воздушного охлаждения мотора;
- Коробка передач – одна передняя;
- Глубина вспашки – до 25 сантиметров;
- Ширина обработки – 35/60 сантиметров;
- Масса агрегата – 48 килограмм.
Небольшой вес модели делает её очень манёвренной, что особенно актуально на участках с преградами. Запчасти к мотоблоку имеют доступную цену, их можно приобрести и установить своими руками.
Крот МК-1А 01 экономично расходует топливо – около одного литра за час эксплуатации мотокультиватора. В качестве топлива лучше использовать марку А-76, масла МГ-8А или М-12 ТП.
Чтобы использовать технику максимально эффективно, понадобиться инструкция к использованию агрегата. Она знакомит с особенностями применения и ухода за культиваторами марки Крот.
Мотокультиватор Крот МК 1А 02
Модель Крот МК 1А 02 выпустили на рынок около 30 лет назад и успела завоевать популярность среди многих фермеров. Сегодня её можно считать устаревшей, но, выполняя ремонт и модернизируя устройство, владельцы смогли сделать культиватор более мощным и функциональным.
Сегодня её можно считать устаревшей, но, выполняя ремонт и модернизируя устройство, владельцы смогли сделать культиватор более мощным и функциональным.
Владельцам небольших земельных участков не выгодно покупать дорогостоящие агрегаты. С работой вполне справиться небольшой мотокультиватор с достаточным уровнем мощности.
Технические параметры модели:
- Двухтактный бензиновый двигатель с мощностью 2,6 л. с.;
- Наличие воздушного охлаждения;
- Коробка передач – одна передняя;
- Вместительность бензобака – 1,8 литра;
- Вес агрегата – 48 килограмм.
Производитель предлагает новые и усовершенствованные модели, но классические агрегаты остаются актуальными из-за своей доступной стоимости. Это отличный помощник для выполнения многих базовых операций в сельском хозяйстве.
Материальные свойства гиалуроновой кислоты голого землекопа
Abstract
Гиалуронан (ГК) является ключевым компонентом внеклеточного матрикса. Учитывая фундаментальную роль ГК в устойчивости голого землекопа к раку (ЯМР), мы предприняли исследование структурных и мягких свойств этого видоспецифического варианта, что является необходимым шагом для его разработки в качестве биоматериала. Исследовали ГК, выделенную из ЯМР головного мозга, легких и кожи, а также выделенную из среды иммортализованных клеток. Как и мышиный HA, NMR HA образует ряд ансамблей, соответствующих широкому распределению молекулярной массы. Однако уникальными для ЯМР являются сильно свернутые структуры, характерная морфология которых зависит от типа ткани. Кожа HA образует плотно упакованные сборки, которые обладают пружинными механическими свойствами в дополнение к сильному сродству к воде. HA головного мозга образует трехмерные складчатые структуры, похожие на макроскопический вид извилин и борозд человеческого мозга. ГК легких образует непроницаемую сеть переплетенных складок, морфология которых может быть описана только как сходство со снежным человеком.
Учитывая фундаментальную роль ГК в устойчивости голого землекопа к раку (ЯМР), мы предприняли исследование структурных и мягких свойств этого видоспецифического варианта, что является необходимым шагом для его разработки в качестве биоматериала. Исследовали ГК, выделенную из ЯМР головного мозга, легких и кожи, а также выделенную из среды иммортализованных клеток. Как и мышиный HA, NMR HA образует ряд ансамблей, соответствующих широкому распределению молекулярной массы. Однако уникальными для ЯМР являются сильно свернутые структуры, характерная морфология которых зависит от типа ткани. Кожа HA образует плотно упакованные сборки, которые обладают пружинными механическими свойствами в дополнение к сильному сродству к воде. HA головного мозга образует трехмерные складчатые структуры, похожие на макроскопический вид извилин и борозд человеческого мозга. ГК легких образует непроницаемую сеть переплетенных складок, морфология которых может быть описана только как сходство со снежным человеком. В отличие от коммерчески доступной ГК, ГК ЯМР легко образует прочные гели без необходимости химического сшивания. Гели ЯМР ГА резко переходят от вязкоупругих к эластичноподобным свойствам при обезвоживании или многократном нагружении. Кроме того, ЯМР-ГА может образовывать упорядоченные тонкие пленки с лежащей в их основе полукристаллической структурой. Учитывая роль ГК в поддержании гидратации кожи, вполне вероятно, что складчатые структуры способствуют как эластичности, так и молодости кожи ЯМР. Также возможно, что такие плотно сложенные материалы могут представлять собой значительный барьер для проникновения клеток в ткани, что является полезной характеристикой для биоматериала.
В отличие от коммерчески доступной ГК, ГК ЯМР легко образует прочные гели без необходимости химического сшивания. Гели ЯМР ГА резко переходят от вязкоупругих к эластичноподобным свойствам при обезвоживании или многократном нагружении. Кроме того, ЯМР-ГА может образовывать упорядоченные тонкие пленки с лежащей в их основе полукристаллической структурой. Учитывая роль ГК в поддержании гидратации кожи, вполне вероятно, что складчатые структуры способствуют как эластичности, так и молодости кожи ЯМР. Также возможно, что такие плотно сложенные материалы могут представлять собой значительный барьер для проникновения клеток в ткани, что является полезной характеристикой для биоматериала.
Введение
Гиалуронан (HA), также известный как 1,2 как гиалуроновая кислота, представляет собой полимер внеклеточного матрикса (ECM), присутствующий в большинстве тканей. В синтезе ГК участвуют три ключевых фермента: ГК-синтаза 1, 2 и 3 (HAS1-3), после чего она расщепляется гиалуронидазой 3 . Регуляция оборота ГК очень важна, поскольку существует четкая связь между метаболизмом ГК и прогрессированием рака, а обилие ГК является показателем плохого прогноза для ряда опухолей у пациентов 4,5,6,7,8 . Роль HA в прогрессировании заболевания, возможно, неудивительна, учитывая ее функцию во взаимодействиях клетка-ECM 9,10,11 . Основным рецептором HA является CD44, рецептор адгезии на клеточной поверхности, который связывает ряд лигандов и сам связан с метастазированием 12 . Биологическая функция ГК часто связана с ее молекулярной массой, при этом диапазон размеров полимера определяется в зависимости от ткани происхождения 13,14 . Учитывая важность взаимодействий ECM-клеток в прогрессировании заболевания и, в частности, взаимодействия HA-клеток, было бы интересно изучить структуру/сборку HA у устойчивых к раку видов.
Регуляция оборота ГК очень важна, поскольку существует четкая связь между метаболизмом ГК и прогрессированием рака, а обилие ГК является показателем плохого прогноза для ряда опухолей у пациентов 4,5,6,7,8 . Роль HA в прогрессировании заболевания, возможно, неудивительна, учитывая ее функцию во взаимодействиях клетка-ECM 9,10,11 . Основным рецептором HA является CD44, рецептор адгезии на клеточной поверхности, который связывает ряд лигандов и сам связан с метастазированием 12 . Биологическая функция ГК часто связана с ее молекулярной массой, при этом диапазон размеров полимера определяется в зависимости от ткани происхождения 13,14 . Учитывая важность взаимодействий ECM-клеток в прогрессировании заболевания и, в частности, взаимодействия HA-клеток, было бы интересно изучить структуру/сборку HA у устойчивых к раку видов.
Голый землекоп (ЯМР, Heterocephalus glaber ) имеет замечательную биологию. Они могут жить в десять раз дольше, чем грызуны того же размера, и у них редко развивается рак 15,16,17 . В 2013 году в статье сообщалось, что ГК способствовала устойчивости к раку ЯМР 18 . Более того, авторы сообщили, что, возможно, форма ГК со сверхвысокой молекулярной массой (более 6100 кДа), обнаруженная в ЯМР, способствовала эластичности его характерной морщинистой кожи. Учитывая такие необычные функции, приписываемые ЯМР ГА, возникает вопрос, есть ли что-нибудь в его структуре или свойствах мягкой материи, что могло бы объяснить его функциональность?
В 2013 году в статье сообщалось, что ГК способствовала устойчивости к раку ЯМР 18 . Более того, авторы сообщили, что, возможно, форма ГК со сверхвысокой молекулярной массой (более 6100 кДа), обнаруженная в ЯМР, способствовала эластичности его характерной морщинистой кожи. Учитывая такие необычные функции, приписываемые ЯМР ГА, возникает вопрос, есть ли что-нибудь в его структуре или свойствах мягкой материи, что могло бы объяснить его функциональность?
Большинство данных о структуре/сборке ГК почти исключительно получены из материала, полученного в результате ферментации бактерий (имеется в продаже) 19,20 . Реже его получают из петушиных гребней или человеческих жидкостей и тканей. Независимо от того, визуализируется ли он в водной среде или на воздухе, обнаружено, что HA образует плоские разветвленные сети волокон 21 . Скотт и др. . использовали ротационную теневую электронную микроскопию, чтобы показать, что ГК (выделенная из петушиных гребней, Streptococci и мезотелиомная жидкость) образует плоские сети в растворе, и чем длиннее молекула ГК, тем больше разветвлений происходит 22 . В более поздних исследованиях использовалась атомно-силовая микроскопия (АСМ) в воздухе (в отличие от водного раствора) для изучения морфологии наноструктур молекулярного масштаба. Наблюдалась сборка как рыхлых клубков отдельных молекулярных цепей, так и протяженных структур с характерными плоскими разветвленными сетевыми структурами, общими 19,21,23,24 . С помощью силовой спектроскопии на основе АСМ удалось развернуть и растянуть одиночные цепи ГК и подтвердить сетевые структуры 25 . В макроскопическом масштабе и изолированно ГК образует только слабый гель и должна быть либо химически сшита, либо объединена в композит, чтобы его можно было использовать в качестве биоматериала 26 .
В более поздних исследованиях использовалась атомно-силовая микроскопия (АСМ) в воздухе (в отличие от водного раствора) для изучения морфологии наноструктур молекулярного масштаба. Наблюдалась сборка как рыхлых клубков отдельных молекулярных цепей, так и протяженных структур с характерными плоскими разветвленными сетевыми структурами, общими 19,21,23,24 . С помощью силовой спектроскопии на основе АСМ удалось развернуть и растянуть одиночные цепи ГК и подтвердить сетевые структуры 25 . В макроскопическом масштабе и изолированно ГК образует только слабый гель и должна быть либо химически сшита, либо объединена в композит, чтобы его можно было использовать в качестве биоматериала 26 .
Хорошо известно, что структурные изменения ГК с течением времени имеют основополагающее значение для старения кожи человека, а ее сродство к воде считается критически важным для поддержания эластичности кожи 27 . На сегодняшний день не было попыток изучить структуру или свойства мягкого вещества ГК с помощью ЯМР. Такое понимание свойств материала ЯМР ГА является предпосылкой для его разработки в качестве потенциального биоматериала для лечения рака. Таким образом, цель этой работы заключалась в очистке ЯМР-ГА и изучении свойств материала и структуры с особым акцентом на различия и сходства между ГК других видов.
Такое понимание свойств материала ЯМР ГА является предпосылкой для его разработки в качестве потенциального биоматериала для лечения рака. Таким образом, цель этой работы заключалась в очистке ЯМР-ГА и изучении свойств материала и структуры с особым акцентом на различия и сходства между ГК других видов.
Результаты и обсуждение
Первой исследованной тканью была кожа, так как характерная морщинистая, но эластичная кожа ЯМР была предположительно приписана присутствию высокомолекулярного HA 18 . Используя биотинилированную версию очень специфичного и прочно связывающегося гиалуронан-связывающего белка (HABP) 28 , гистологический анализ подошвенной поверхности кожи передней лапы показал, что HA присутствует как в мышиной, так и в ЯМР-коже. Однако ГК наблюдался в гораздо большей степени в эпидермисе ЯМР по сравнению с мышами, где, как показали другие исследования, 29 , HA в основном ограничивался дермой (рис. 1a,b). ГК, экстрагированная из культуральной среды иммортализованных фибробластов кожи методом ЯМР, представляла ряд собранных конформаций, в результате чего можно было четко разделить отдельные цепи (рис. 1c, d).
1c, d).
ЯМР кожи HA. ( a ) Передняя лапа мыши, подошвенный срез кожи, окрашенный на HA с использованием HABP и стрептавидина-Alexa488 (зеленый) и окрашивание ядер DAPI (синий). ( b ) ЯМР передней лапы, подошвенный срез кожи, окрашенный на HA с использованием HABP и стрептавидина-Alexa488 (зеленый) и окрашивание ядер DAPI (синий). В отличие от мыши, ЯМР HA распространяется глубоко в эпидермис и значительно более обильна. ( c ) Изображение топографии АСМ цепей ГК, полученных из ГК, экстрагированных из среды ЯМР фибробластов кожи. ( d ) Топографическое изображение АСМ другого типа сборки цепи ГК из фибробластов кожи ЯМР.
Изображение в натуральную величину
Размеры этих цепочек соответствовали пучку из нескольких полимерных молекул со средней шириной 33,6 ± 7,71 нм (n = 100) и высотой, в несколько раз превышающей пару нанометров, характерную высоту отдельные молекулы. Наиболее распространенными структурами в образце были четко определенные, плотно упакованные суперспирали (рис. 2а, б). Такие плотно свернутые объекты, по-видимому, являются основной единицей ГК в коже ЯМР с характерным видом «цветной капусты» при визуализации с помощью СЭМ (рис. 2c). ЯМР ГК, экстрагированная из ткани кожи (а не из среды культивируемых фибробластов кожи), также образовывала такие же сверхскрученные складчатые структуры, хотя в целом они были крупнее, чем у ГК, экстрагированной из культуральной среды (рис. 2d,e). Сети отдельных цепей также можно наблюдать для образцов, извлеченных из ткани кожи ЯМР (рис. 2f), демонстрируя согласованность между ГК, извлеченной из клеточной среды, и ГК, извлеченной из ткани. Используя конический наконечник АСМ, можно было адресовать отдельные суперспирали и определить их свойства мягкого вещества с помощью вдавливания. Значения модуля Юнга можно получить, подгоняя кривую вдавливания к модели Снеддона, уравнение 1 9{2} $$
2а, б). Такие плотно свернутые объекты, по-видимому, являются основной единицей ГК в коже ЯМР с характерным видом «цветной капусты» при визуализации с помощью СЭМ (рис. 2c). ЯМР ГК, экстрагированная из ткани кожи (а не из среды культивируемых фибробластов кожи), также образовывала такие же сверхскрученные складчатые структуры, хотя в целом они были крупнее, чем у ГК, экстрагированной из культуральной среды (рис. 2d,e). Сети отдельных цепей также можно наблюдать для образцов, извлеченных из ткани кожи ЯМР (рис. 2f), демонстрируя согласованность между ГК, извлеченной из клеточной среды, и ГК, извлеченной из ткани. Используя конический наконечник АСМ, можно было адресовать отдельные суперспирали и определить их свойства мягкого вещества с помощью вдавливания. Значения модуля Юнга можно получить, подгоняя кривую вдавливания к модели Снеддона, уравнение 1 9{2} $$
(1)
Где:
F Sneddon = Force
E Surface = Модуль Янга на поверхности
v Surface = Коэффициент поверхности Поэссон. угол раскрытия конуса
угол раскрытия конуса
с 0 = точка нулевого отпечатка
с 0 - с = глубина отпечатка
Рисунок 2ЯМР кожи ГА. ( a ) Сканирующая электронная микроскопия (СЭМ) ЯМР HA, извлеченного из среды фибробластов кожи ЯМР. ( b ) Сверхвитки, связанные между собой большим волокном. ( c ) СЭМ высокого разрешения, показывающая высокую плотность складок ( d ) АСМ ГК, выделенной из ЯМР-среды фибробластов кожи ( e ) АСМ ЯМР ГК, выделенной из кожной ткани. снова демонстрируя высокую степень складки, как в случае HA, экстрагированного из фибробластов ЯМР. ( f ) АСМ отдельных цепей ГК, выделенных из ткани кожи методом ЯМР.
Полноразмерное изображение
На рис. 3а показано топографическое изображение отдельной суперспирали NMRHA, полученное с помощью АСМ, а на рис. 3b показаны отдельные положения в суперспирале, где были сделаны отдельные измерения вдавливания (зеленые квадраты). Типичная кривая сила-разделение для ЯМР ГК, извлеченной из кожи, представлена на рис. 3c. Особого внимания заслуживают пики на красной кривой ретракции, которые представляют собой развертывание полимерных цепей. Это не однородный пилообразный рисунок, а скорее наблюдается неравномерность ширины пиков, вероятно, отражающая складчатые домены разного размера. Необычной особенностью этого паттерна являются пики в профиле вдавливания, показанные синим цветом, что можно интерпретировать как развертывание структуры при сжатии. Из этих кривых вдавливания внутри суперспиралей ГК ЯМР кожи было получено значение модуля Юнга, которое составило 13,77 ± 2,44 МПа (рис. 3d).
Типичная кривая сила-разделение для ЯМР ГК, извлеченной из кожи, представлена на рис. 3c. Особого внимания заслуживают пики на красной кривой ретракции, которые представляют собой развертывание полимерных цепей. Это не однородный пилообразный рисунок, а скорее наблюдается неравномерность ширины пиков, вероятно, отражающая складчатые домены разного размера. Необычной особенностью этого паттерна являются пики в профиле вдавливания, показанные синим цветом, что можно интерпретировать как развертывание структуры при сжатии. Из этих кривых вдавливания внутри суперспиралей ГК ЯМР кожи было получено значение модуля Юнга, которое составило 13,77 ± 2,44 МПа (рис. 3d).
Для сравнения, ГК экстрагировали и очищали из ткани кожи человека. В отличие от ГК кожи ЯМР, в ГК кожи человека не было сверхскрученных структур, но вместо этого наблюдались большие плоские сети, как показано с помощью СЭМ (рис. 4а) и АСМ (рис. 4б). Профили вдавливания были типичными для мягкого эластичного материала (рис. 4с) без разворачивающихся пиков ни на кривой приближения, ни на кривой ретракции. Путем обращения к наконечнику АСМ по всей сети было определено, что модуль Юнга составляет 820,00 МПа ± 14,00 МПа, что значительно жестче, чем ЯМР HA в форме суперспирали.
4с) без разворачивающихся пиков ни на кривой приближения, ни на кривой ретракции. Путем обращения к наконечнику АСМ по всей сети было определено, что модуль Юнга составляет 820,00 МПа ± 14,00 МПа, что значительно жестче, чем ЯМР HA в форме суперспирали.
НА также экстрагировали из ЯМР головного мозга и легочной ткани. ГК, извлеченная из целого мозга, образовывала большие (диаметром не менее нескольких микрон) суперспирали, но на этот раз складки были менее плотными и напоминали макроскопический вид извилин и борозд головного мозга человека (рис. 5а, б). ГК, извлеченная из легкого с помощью ЯМР, также образовывала суперспирали, но на этот раз характерная складчатая структура напоминала морфологию, которую можно описать только как напоминающую снеговика (рис. 5c). Изображения с более высоким разрешением показали, что эти необычные морфологии состоят из сети переплетенных цепей на высокая плотность (рис. 5d). Структуры ГА головного мозга и легких были очень плотно сложены с несколькими видимыми промежутками. Это резко контрастировало с ГК, экстрагированной из кожи человека, в которой суперспирали не образовывались, а случайные сети цепей были доминирующими структурами. Коммерчески доступная ГК (высокомолекулярная, полученная путем микробной ферментации Streptococcus pyogenes , R&D systems) образовали разветвленные сети.
Это резко контрастировало с ГК, экстрагированной из кожи человека, в которой суперспирали не образовывались, а случайные сети цепей были доминирующими структурами. Коммерчески доступная ГК (высокомолекулярная, полученная путем микробной ферментации Streptococcus pyogenes , R&D systems) образовали разветвленные сети.
Профили вдавливания с пиками на кривой вдавливания были видны только для суперспиралей кожи, как на рис. 3c. При погружении суперспиралей легкого ГА в воду (комнатной температуры) происходит изменение их механических свойств. После 10 минут погружения в воду суперзмеевик сохраняет свои упругие характеристики (рис. 5д), но затем через 20 минут кривая отрыва сил принимает характерный вид вязкоупругого материала (рис. 5е). Это имело место для ГК из всех исследованных ЯМР тканей/клеток. Для точного измерения модуля Юнга суперспиралей до и после гидратации использовали сферический зонд. Переход между вязкоупругим и упругим, то есть от рис. 5f к рис. 5e, может быть получен повторным вдавливанием в одно и то же положение. Это многократное нагружение (иногда до 150 вдавливаний с усилием до 50 нН) как бы выдавливает воду. Переход внезапный с изменением формы кривой силового расстояния между двумя последовательными углублениями. Чисто эластичное состояние также может быть достигнуто путем обезвоживания ГК.
Это многократное нагружение (иногда до 150 вдавливаний с усилием до 50 нН) как бы выдавливает воду. Переход внезапный с изменением формы кривой силового расстояния между двумя последовательными углублениями. Чисто эластичное состояние также может быть достигнуто путем обезвоживания ГК.
Для ГК, экстрагированной из всех тканей ЯМР, т. е. головного мозга, легких и кожи, спонтанно образуются прочные макроскопические гели, если оставить 30 мкл раствора ГК (экстракт ГК в воде) на поверхности слюды на 15 минут для высыхания, оставляя круглую гель диаметром до 1 см (рис. 6а). ГК, экстрагированная из культуральной среды иммортализованных фибробластов кожи методом ЯМР, демонстрировала такое же гелеобразующее поведение, но такого гелеобразования не наблюдалось для ГК из кожи мыши или человека. По мере обезвоживания геля отчетливо видны составляющие его микрогели (рис. 6b). Гели имеют четко очерченные границы (рис. 6c, d), а составляющие суперспирали восстанавливаются при последующем обезвоживании. Кривые вдавливания частиц геля были аппроксимированы моделью Герца для сфер (рис. 6e) на плоской поверхности, которая предполагает, что глубина вдавливания намного меньше диаметра зонда. Используя сферический зонд и подгоняя кривую вдавливания к модели Герца, ур. 2, определяли модуль Юнга микрогелей. 9{3/2} $$
Кривые вдавливания частиц геля были аппроксимированы моделью Герца для сфер (рис. 6e) на плоской поверхности, которая предполагает, что глубина вдавливания намного меньше диаметра зонда. Используя сферический зонд и подгоняя кривую вдавливания к модели Герца, ур. 2, определяли модуль Юнга микрогелей. 9{3/2} $$
(2)
Где:
F HERZ = FIRCH
E Surface = Модуль Янга на поверхности
V Surface = Ratio Surface Poasson’s Ratio R . TIP = Радиус шарика S 0 = точка нуля вдавливания S 0 -S = глубина отступления ( A ) AFM Topography Image of SuperCoiled NRM Struftra. Это сильно складчатые структуры до микрона в высоту и с некоторыми складками/изгибами в полмикрона в диаметре. ( b ) То же изображение с наложенной картой сил. Зеленые точки указывают, где проводятся измерения отдельных отпечатков. Полноразмерное изображение Это были 10,21 ± 1,68 кПа, 10,44 ± 1,31 кПа, 7,86 ± 0,84 кПа для ЯМР кожи, головного мозга и ткани легких соответственно. Кожа человека HA. ( a ) СЭМ выявляет большие плоские сети с четко очерченными отверстиями диаметром до нескольких микрон. ( b ) АСМ, выявляющая ту же форму ГА, что соответствует изображениям СЭМ. ( c ) Кривая сила-разделение, показывающая вдавливание в сеть ГК кожи человека, демонстрирующую упругое поведение. ( d ) Гистограмма значений модуля Юнга для углублений в коже человека HA, полученная в больших сетях. Кривые соответствуют модели Снеддона, что дает среднее значение модуля Юнга 820,00 МПа ± 14,00 МПа для 3000 кривых. Полноразмерное изображение ЯМР ГК, извлеченная из головного мозга и легких. Изображение в полный размер ( a ) Макроскопический гель, образованный ЯМР ГК, извлеченной из кожной ткани. Изображение с полным размером Мышиный ГК (полученный из среды фибробластов кожи) не образует суперспиралей, хотя существуют менее свернутые сферические частицы (рис. 7а). Наиболее распространенной наблюдаемой морфологией являются плотные крупномасштабные сети, в которых отдельные волокна не могут быть разрешены (рис. HA, экстрагированный из среды фибробластов кожи мыши. ( a ) СЭМ показывает типичную сферическую частицу без суперскрученной структуры. ( b ) СЭМ показывает мышиные волокна ГК, которые формируются с большей вероятностью, чем для образцов ЯМР. ( c , d ) АСМ топографические изображения случайных сетей HA мыши. ( e ) SEM высокого разрешения мышиного HA волокна. Изображение в полный размер Хотя суперспирали являются доминирующей морфологией для ЯМР ГА (рис. 8a,b), иногда образуются волокна. Механические свойства, определяемые индентированием редкого волокна, которое можно найти (рис. 8c), более жесткие, чем у супервитков, хотя механическая неоднородность вдоль волокна очевидна из распределения значений модулей. Волокна ГК мыши кажутся более мягкими, но абсолютные значения значений модуля с использованием конических наконечников могут быть ненадежными из-за неопределенностей в области контакта между наконечником и образцом. Изображение в натуральную величину При более высоких концентрациях адсорбированный NMRHA покрывает поверхность и образует тонкую пленку толщиной до нескольких микрон. Иногда пленка сворачивается сама по себе, что дает возможность наблюдать ее внутреннюю структуру (рис. 9а) и определять ее толщину до 1 микрона. При более высоком разрешении разрешается ламеллярная структура, то есть регулярно расположенные листы, в данном случае сложенные попарно (рис. 9b). Структура внутри каждой стопки состоит из многократно свернутых цепочек (рис. Тонкая пленка гиалуроновой кислоты с голым землекопом. ( a ) Топографическое изображение АСМ, на котором тонкая пленка сложена сама по себе. Выявлена толщина пленки порядка микрона. ( b ) Топографическое изображение АСМ, показывающее ламеллярную структуру тонкой пленки. Выступы, состоящие из упорядоченных полимерных цепей, уложены попарно, каждый с одинаковым межплоскостным расстоянием и морфологией. ( c ) Цепи, сложенные вдоль ламеллярной тонкой пленки ЯМР ГА. Полноразмерное изображение Неспособность коммерчески доступной ГК самопроизвольно образовывать прочные гели широко описана в литературе, при этом химическое сшивание или включение в композит является необходимостью для практического использования в качестве биоматериала 26,30, 31 . Одним из примеров, когда это происходит, являются амилоидные фибриллы, встречающиеся в природных клеях. Такая способность упруго деформироваться в нескольких направлениях была бы предпочтительной для материала, который подвергается сдвиговым усилиям, таким как те, которые испытывает кожа, когда животное сжимает и скручивает однопометников в туннелях. Ясно, что микронная морфология ЯМР HA связана с тканью происхождения. Как возникают эти различные морфологии, лучше всего понять с точки зрения их наноразмерной иерархической архитектуры. Мы можем считать базовым звеном свернутую полимерную цепь, и именно степень складчатости определяет морфологию окончательной сборки. В случае ГК легкого ЯМР цепи плотно свернуты (рис. 5d), что приводит к плотной морфологии микронного масштаба без пор (рис. 5c). Сравните это с менее плотно упакованными полимерными цепями (рис. В случае ЯМР вполне вероятно, что сверхвысокая молекулярная масса вносит свой вклад в упругие свойства кожи посредством сверхспирализации. Кроме того, увеличенная площадь поверхности открытой ГК улучшит водоудерживающую способность, что является еще одним важным фактором антивозрастных свойств кожи ЯМР. Эта способность улавливать воду подтверждается образованием микрогелей с поперечными связями, обычно необходимыми для прочных гелей, замененными самовзаимодействиями через запутанный полимер; важно, что эти микрогели не встречаются в ГК, экстрагированной из кожи мыши или человека. Именно эти уникальные водоудерживающие свойства ГК, скорее всего, способствуют необычным свойствам ЯМР-кожи, а не низкой жесткости ЯМР-ГК по сравнению с ГК кожи человека. Это связано с тем, что ГК не является основным структурным компонентом кожи, а коллаген и эластин, скорее всего, способствуют свойствам растяжения/сжатия. Все эксперименты на животных проводились в соответствии с Законом Соединенного Королевства о животных (научные процедуры) 1986 г. с поправками 2012 г. в соответствии с лицензией на проект (70/7705), выданной E. St. J. S. Министерством здравоохранения Офис; Орган Кембриджского университета по этике защиты животных также утвердил процедуры. В этом исследовании использовали ЯМР молодых взрослых (как мужчин, так и женщин). Животных содержали в изготовленной на заказ системе клеток с обычными клетками для мышей/крыс, соединенными туннелем разной длины. Образцы кожи собирали с подушечек лап 6-8-недельных самок мышей (n = 2) и ЯМР (n = 2), постфиксировали в 4% параформальдегиде (PFA) в течение 1 ч и инкубировали в течение ночи при 4 °C в 30% сахарозе (масса/объем) для криозащиты. Образцы кожи помещали (эпидермальной стороной вверх) в матрицу для заливки Shandon M-1 (Thermo Fisher Scientific), быстро замораживали в 2-метилбутане (Honeywell International) на сухом льду и хранили при температуре -80 °C до дальнейшей обработки. После умерщвления животного брали кожу из области подмышек или нижней части живота, очищали от жира или мышечной ткани, обильно обрызгивали 70%-ным этанолом и мелко измельчали стерильными скальпелями. Измельченную кожу затем смешивали с 5 мл среды для изоляции клеток ЯМР (DMEM с высоким содержанием глюкозы (Gibco № 11965092) с добавлением 100 ед. смеси ферментов клеточной диссоциации (10 мг мл -1 Коллагеназа (Roche # 110887), 1000 ЕД мл -1 Гиалуронидаза (Sigma # h4506) в среде DMEM с высоким содержанием глюкозы (Gibco # 11965092)) и инкубировали при 37°C в течение 3–5 часов. После декапитации ткани удаляли, взвешивали и хранили при -80 °C до проведения экстракции ГК. Ткани расщепляли при 50°C в течение ночи в буфере для расщепления (10 мМ Tris-CL, 25 мМ EDTA, 100 мМ NaCl, 0,5% SDS и 0,1 мкг/мл протеиназы K). На следующий день образцы центрифугировали при 18 000×g в течение 10 минут для удаления остаточных частиц ткани, а супернатант переносили в новую пробирку. В супернатант добавляли четыре объема этанола с последующей инкубацией в течение ночи при -20 °С. Иммортализованные фибробласты кожи высевали и выдерживали в течение 7 дней без смены среды. Через 7 дней среду из каждой колбы собирали и центрифугировали при 4000×g в течение 10 минут для удаления остаточных клеток. Этическое разрешение на получение кожи человека получено от Комитета по этике исследований, Северо-Восточный Ньюкасл 1, номер ссылки: REC 12/NE/0395. После информированного согласия образцы кожи были получены у пациентов, перенесших ортопедическую операцию. Все эксперименты проводились в соответствии с положениями Закона Соединенного Королевства о тканях человека (2004 г. В каждой точке можно выполнить несколько измерений, а точный контроль кантилевера достигается с помощью сканера с обратной связью. ( c ) Кривая разделения сил (пирамидальный наконечник) для суперспирали HA кожи ЯМР, измеренная в воздухе. Кривая ретракции (красная) кончика АСМ показывает несколько пиков на микронной длине, что свидетельствует о развертывании больших цепей ГК. Что необычно для кривой разделения усилий, кривая приближения (синяя) также демонстрирует несколько пиков силы. Мы приписываем это силе, необходимой для того, чтобы оттолкнуть сверхскрученные цепи в сторону, когда игла проникает сквозь структуру. ( d ) Гистограмма значений модуля Юнга для вдавливания в суперспирали ГА кожи ЯМР, полученная для 5 катушек со 112 кривыми, соответствующими модели Снеддона, дает среднее значение модуля Юнга 13,67 МПа ± 2,44 МПа.
В каждой точке можно выполнить несколько измерений, а точный контроль кантилевера достигается с помощью сканера с обратной связью. ( c ) Кривая разделения сил (пирамидальный наконечник) для суперспирали HA кожи ЯМР, измеренная в воздухе. Кривая ретракции (красная) кончика АСМ показывает несколько пиков на микронной длине, что свидетельствует о развертывании больших цепей ГК. Что необычно для кривой разделения усилий, кривая приближения (синяя) также демонстрирует несколько пиков силы. Мы приписываем это силе, необходимой для того, чтобы оттолкнуть сверхскрученные цепи в сторону, когда игла проникает сквозь структуру. ( d ) Гистограмма значений модуля Юнга для вдавливания в суперспирали ГА кожи ЯМР, полученная для 5 катушек со 112 кривыми, соответствующими модели Снеддона, дает среднее значение модуля Юнга 13,67 МПа ± 2,44 МПа. Типичное распределение модуля, взятое из почти 1000 кривых вдавливания, показано на рис. 6f. Эластичные суперспирали (после обезвоживания) обладают гораздо лучшими механическими свойствами, чем микрогели, например, суперспирали ЯМР мозга имеют модуль Юнга 30,31 ± 1,54 МПа (n = 149).9).
Типичное распределение модуля, взятое из почти 1000 кривых вдавливания, показано на рис. 6f. Эластичные суперспирали (после обезвоживания) обладают гораздо лучшими механическими свойствами, чем микрогели, например, суперспирали ЯМР мозга имеют модуль Юнга 30,31 ± 1,54 МПа (n = 149).9). ( a ) АСМ Топографическое изображение свернутой ЯМР ГК, выделенной из целого мозга ( b ) Топографическое АСМ изображение высокого разрешения, демонстрирующее характерные «мозгоподобные» складки ГК, выделенной из всего мозга ЯМР. ( c ) ГК, извлеченная из легких с помощью ЯМР, образует характерные структуры «снежного человека». ( d ) Сканирование HA с более высоким разрешением с помощью ЯМР легкого, где можно разрешить плотно свернутые цепи, образующие непроницаемую сеть. ( e ) Кривая отступа для ЯМР легкого HA, с отступом несколько раз (более 100) в одном и том же месте. Вода была выдавлена, и теперь суперспираль имеет характеристики чисто эластичного материала. ( f ) Кривая вдавливания для ГК легких ЯМР после гидратации в течение часа. Он имеет характерную для гидрогеля форму.
( a ) АСМ Топографическое изображение свернутой ЯМР ГК, выделенной из целого мозга ( b ) Топографическое АСМ изображение высокого разрешения, демонстрирующее характерные «мозгоподобные» складки ГК, выделенной из всего мозга ЯМР. ( c ) ГК, извлеченная из легких с помощью ЯМР, образует характерные структуры «снежного человека». ( d ) Сканирование HA с более высоким разрешением с помощью ЯМР легкого, где можно разрешить плотно свернутые цепи, образующие непроницаемую сеть. ( e ) Кривая отступа для ЯМР легкого HA, с отступом несколько раз (более 100) в одном и том же месте. Вода была выдавлена, и теперь суперспираль имеет характеристики чисто эластичного материала. ( f ) Кривая вдавливания для ГК легких ЯМР после гидратации в течение часа. Он имеет характерную для гидрогеля форму. ( b ) Край макроскопического геля, сформированного из головного мозга ЯМР HA. ( c ) Частицы геля ГК ЯМР из (полученного из ткани головного мозга) ( d ) Край макроскопического геля, образованного из ГК ЯМР легких. ( e ) Кривая вдавливания геля, полученного из ЯМР кожи ГК. Для проведения этих измерений использовались сферические зонды, а модель Герца для сферы, вдавливающейся в поверхность, была подогнана к кривой вдавливания для извлечения модуля Юнга. ( f ) Гистограмма значений модуля Юнга для вдавливания в частицы геля ГК легких ЯМР с использованием сферических зондов. Среднее значение модуля Юнга 7,86 кПа ± 0,83 кПа для 999 кривых.
( b ) Край макроскопического геля, сформированного из головного мозга ЯМР HA. ( c ) Частицы геля ГК ЯМР из (полученного из ткани головного мозга) ( d ) Край макроскопического геля, образованного из ГК ЯМР легких. ( e ) Кривая вдавливания геля, полученного из ЯМР кожи ГК. Для проведения этих измерений использовались сферические зонды, а модель Герца для сферы, вдавливающейся в поверхность, была подогнана к кривой вдавливания для извлечения модуля Юнга. ( f ) Гистограмма значений модуля Юнга для вдавливания в частицы геля ГК легких ЯМР с использованием сферических зондов. Среднее значение модуля Юнга 7,86 кПа ± 0,83 кПа для 999 кривых. 7b, c). Отдельные волокна более многочисленны у образцов мышей (рис. 7d, e).
7b, c). Отдельные волокна более многочисленны у образцов мышей (рис. 7d, e). Ясно то, что волокна ЯМР жестче, чем суперспирали. 9Рисунок 8 ( b ) СЭМ с большим полем, показывающая изобилие / покрытие поверхности суперспиралей ЯМР HA. ( c ) Гистограмма значений модуля Юнга, полученных из волокна ЯМР HA со средним значением 0,28 ГПа ± 0,0165 ГПа для 100 кривых. ( d ) SEM ЯМР-волокна HA (e) Гистограмма значений модуля Юнга, взятых из волокна HA мыши, со средним значением 78,025 MPa ± 7,533 MPa для 100 кривых.
Ясно то, что волокна ЯМР жестче, чем суперспирали. 9Рисунок 8 ( b ) СЭМ с большим полем, показывающая изобилие / покрытие поверхности суперспиралей ЯМР HA. ( c ) Гистограмма значений модуля Юнга, полученных из волокна ЯМР HA со средним значением 0,28 ГПа ± 0,0165 ГПа для 100 кривых. ( d ) SEM ЯМР-волокна HA (e) Гистограмма значений модуля Юнга, взятых из волокна HA мыши, со средним значением 78,025 MPa ± 7,533 MPa для 100 кривых. 9).в) таких же размеров, как в сверхскрученных структурах. Однако пленка не образует гель при гидратации, скорее всего, из-за ее полукристаллической природы, при которой соседние полимерные цепи вынуждены взаимодействовать друг с другом, а не с водой.
9).в) таких же размеров, как в сверхскрученных структурах. Однако пленка не образует гель при гидратации, скорее всего, из-за ее полукристаллической природы, при которой соседние полимерные цепи вынуждены взаимодействовать друг с другом, а не с водой. Выводы
 Напротив, природа фолдинга ЯМР-ГА необычна. Он образует конформацию, подобную шару из резиновых лент, и в случае ГК, извлеченной из кожи, это приводит к уникальному профилю вдавливания. Более того, хотя точную концентрацию для таких малых количеств биополимера определить невозможно, интерес представляет способность ЯМР ГА образовывать гели без необходимости химического сшивания. Был выполнен большой объем работы по вдавливанию и развертыванию мягких биологических веществ, включая белки, волокна, такие как коллаген, клеточные мембраны, липидные бислои и углеводы. Однако редко можно найти материал с пилообразной кривой вдавливания (в отличие от кривой ретракции).
Напротив, природа фолдинга ЯМР-ГА необычна. Он образует конформацию, подобную шару из резиновых лент, и в случае ГК, извлеченной из кожи, это приводит к уникальному профилю вдавливания. Более того, хотя точную концентрацию для таких малых количеств биополимера определить невозможно, интерес представляет способность ЯМР ГА образовывать гели без необходимости химического сшивания. Был выполнен большой объем работы по вдавливанию и развертыванию мягких биологических веществ, включая белки, волокна, такие как коллаген, клеточные мембраны, липидные бислои и углеводы. Однако редко можно найти материал с пилообразной кривой вдавливания (в отличие от кривой ретракции). Учитывая наличие относительно большого количества ГК в коже ЯМР (рис. 1b) 18 , вполне вероятно, что эти суперспирали способствуют ее упругой природе. Эти суперспирали с их характерной структурой, подобной цветной капусте, отсутствуют в ГК, экстрагированной из кожи человека и мыши, а также отсутствуют в коммерчески доступной ГК, продуцируемой бактериями. С точки зрения соотношения структура-функция ГК хорошо известно, что молекулярная масса ГК может быть связана с функцией в исходной ткани 32 .
Учитывая наличие относительно большого количества ГК в коже ЯМР (рис. 1b) 18 , вполне вероятно, что эти суперспирали способствуют ее упругой природе. Эти суперспирали с их характерной структурой, подобной цветной капусте, отсутствуют в ГК, экстрагированной из кожи человека и мыши, а также отсутствуют в коммерчески доступной ГК, продуцируемой бактериями. С точки зрения соотношения структура-функция ГК хорошо известно, что молекулярная масса ГК может быть связана с функцией в исходной ткани 32 . 5b) ГК ЯМР головного мозга, что приводит к более трехмерной сборке микронного масштаба, содержащей поры (рис. 5a). Можно предположить, что именно эти плотно складчатые структуры с небольшим количеством пор размером с клетку могут образовывать непреодолимый барьер для инвазии, необходимой для метастазирования рака и роста опухоли. Однако важно не забывать о той роли, которую гиалуронан играет в клеточной передаче сигналов посредством его взаимодействия с рецептором CD44, связанным с клеточной мембраной, и о том, что раковые клетки расщепляют этот рецептор с использованием матриксных металлопротеиназ. Сверхспиральная структура ЯМР HA может предотвратить такое расщепление. Также вероятно, что именно улучшенные водоудерживающие свойства ГК ЯМР сохраняют «молодость» внеклеточного матрикса так же, как гиалуронан сохраняет кожу увлажненной. Недавно было показано, что пожилой внеклеточный матрикс более восприимчив к инвазии раковых клеток, чем внеклеточный матрикс более молодых людей 33 .
5b) ГК ЯМР головного мозга, что приводит к более трехмерной сборке микронного масштаба, содержащей поры (рис. 5a). Можно предположить, что именно эти плотно складчатые структуры с небольшим количеством пор размером с клетку могут образовывать непреодолимый барьер для инвазии, необходимой для метастазирования рака и роста опухоли. Однако важно не забывать о той роли, которую гиалуронан играет в клеточной передаче сигналов посредством его взаимодействия с рецептором CD44, связанным с клеточной мембраной, и о том, что раковые клетки расщепляют этот рецептор с использованием матриксных металлопротеиназ. Сверхспиральная структура ЯМР HA может предотвратить такое расщепление. Также вероятно, что именно улучшенные водоудерживающие свойства ГК ЯМР сохраняют «молодость» внеклеточного матрикса так же, как гиалуронан сохраняет кожу увлажненной. Недавно было показано, что пожилой внеклеточный матрикс более восприимчив к инвазии раковых клеток, чем внеклеточный матрикс более молодых людей 33 . Таким образом, можно предположить, что внеклеточный матрикс ЯМР проявляет меньше признаков старения, чем другие короткоживущие организмы, и, следовательно, менее восприимчив к инвазивному раку.
Таким образом, можно предположить, что внеклеточный матрикс ЯМР проявляет меньше признаков старения, чем другие короткоживущие организмы, и, следовательно, менее восприимчив к инвазивному раку. Другой важной характеристикой спирального ГА ЯМР является плотность полимерной сетки. Видимых зазоров мало из-за сложенной упаковки. Это особенно поразительно по сравнению с морфологией сетей HA человека и мыши. Имитация суперскрученной структуры ЯМР-ГК в синтетических или искусственных природных полимерах может стать стратегией изучения воссоздания эластичности и омолаживающих эффектов в тканевой инженерии кожи, а также в биоматериалах для предотвращения инвазии клеток.
Другой важной характеристикой спирального ГА ЯМР является плотность полимерной сетки. Видимых зазоров мало из-за сложенной упаковки. Это особенно поразительно по сравнению с морфологией сетей HA человека и мыши. Имитация суперскрученной структуры ЯМР-ГК в синтетических или искусственных природных полимерах может стать стратегией изучения воссоздания эластичности и омолаживающих эффектов в тканевой инженерии кожи, а также в биоматериалах для предотвращения инвазии клеток. Материалы и методы
Животные
 Подстилка и материал для гнезд были предоставлены вместе с беговым колесом. Комнату прогревали до 28 °C с помощью нагревательного кабеля для дополнительного обогрева, проложенного под 2–3 клетками, и использовали красное освещение (08:00–16:00). ЯМР были убиты воздействием возрастающей концентрации CO 2 с последующим обезглавливанием. Мыши C57Bl6J содержались в помещении с контролируемой температурой (21 °C) с 12-часовым циклом свет/темнота, с доступом к пище и воде ad libitum в группах до пяти человек. Для выделения тканей использовали самок мышей не старше 12 недель.
Подстилка и материал для гнезд были предоставлены вместе с беговым колесом. Комнату прогревали до 28 °C с помощью нагревательного кабеля для дополнительного обогрева, проложенного под 2–3 клетками, и использовали красное освещение (08:00–16:00). ЯМР были убиты воздействием возрастающей концентрации CO 2 с последующим обезглавливанием. Мыши C57Bl6J содержались в помещении с контролируемой температурой (21 °C) с 12-часовым циклом свет/темнота, с доступом к пище и воде ad libitum в группах до пяти человек. Для выделения тканей использовали самок мышей не старше 12 недель. Иммуногистохимия ЯМР и образцов кожи мышей
 В день иммунного окрашивания образцы встроенной кожи разрезали на срезы толщиной 12 мкм с помощью криостата Leica (CM3000; Nussloch, Германия) и помещали на предметные стекла Superfrost Plus (Thermo Fisher Scientific). Для иммуногистохимии случайно выбранные предметные стекла дважды промывали PBS-Tween, затем блокировали раствором для разведения антител, состоящим из 0,2% (об./об.) Triton X-100 и 2% (об./об.) бычьего сывороточного альбумина в PBS. , в течение двух часов при комнатной температуре (~22 °C). На предметные стекла добавляли антитело к биотинилированному белку, связывающему гиалуронан (b-HABP) (amsbio, AMS.HKD-BC41, 1:200 в разбавителе антител), и инкубировали в течение ночи при 4 °C. На следующий день предметные стекла трижды промывали с помощью PBS-Tween и инкубировали в течение двух часов при комнатной температуре со стрептавидином Alexa Fluor 488 (Invitrogen, S11223, 1:1000 в PBS). После завершения связывания биотина со стрептавидином предметные стекла трижды промывали PBS-Tween и инкубировали с ядерным красителем DAPI (Sigma, D9).
В день иммунного окрашивания образцы встроенной кожи разрезали на срезы толщиной 12 мкм с помощью криостата Leica (CM3000; Nussloch, Германия) и помещали на предметные стекла Superfrost Plus (Thermo Fisher Scientific). Для иммуногистохимии случайно выбранные предметные стекла дважды промывали PBS-Tween, затем блокировали раствором для разведения антител, состоящим из 0,2% (об./об.) Triton X-100 и 2% (об./об.) бычьего сывороточного альбумина в PBS. , в течение двух часов при комнатной температуре (~22 °C). На предметные стекла добавляли антитело к биотинилированному белку, связывающему гиалуронан (b-HABP) (amsbio, AMS.HKD-BC41, 1:200 в разбавителе антител), и инкубировали в течение ночи при 4 °C. На следующий день предметные стекла трижды промывали с помощью PBS-Tween и инкубировали в течение двух часов при комнатной температуре со стрептавидином Alexa Fluor 488 (Invitrogen, S11223, 1:1000 в PBS). После завершения связывания биотина со стрептавидином предметные стекла трижды промывали PBS-Tween и инкубировали с ядерным красителем DAPI (Sigma, D9). 452, 1:1000 в PBS), в течение 10 мин. Затем слайды один раз промывали PBS-Tween, монтировали и визуализировали с помощью микроскопа Olympus BX51 (Токио, Япония) и камеры QImaging (Суррей, Канада). Все слайды были визуализированы с одинаковыми уровнями экспозиции на каждой длине волны (488 нм для b-HABP и 350 нм для DAPI), и для всех слайдов в ImageJ было сделано одинаковое повышение контрастности. Предметные стекла, которые не инкубировали с конъюгатом стрептавидина (отрицательный контроль), не проявляли флуоресценции.
452, 1:1000 в PBS), в течение 10 мин. Затем слайды один раз промывали PBS-Tween, монтировали и визуализировали с помощью микроскопа Olympus BX51 (Токио, Япония) и камеры QImaging (Суррей, Канада). Все слайды были визуализированы с одинаковыми уровнями экспозиции на каждой длине волны (488 нм для b-HABP и 350 нм для DAPI), и для всех слайдов в ImageJ было сделано одинаковое повышение контрастности. Предметные стекла, которые не инкубировали с конъюгатом стрептавидина (отрицательный контроль), не проявляли флуоресценции. Получение иммортализованных клеточных линий
 Кожу кратко встряхивали каждые 30 минут, чтобы способствовать диссоциации клеток, и вручную проверяли диссоциацию клеток. После полной диссоциации клетки осаждали центрифугированием при 500 g в течение 5 минут и ресуспендировали в ЯМР среде для культивирования клеток (DMEM с высоким содержанием глюкозы (Gibco # 11965092) с добавлением 15% эмбриональной бычьей сыворотки (Gibco), заменимых аминокислот (Gibco # 11965092). 11140050), 1 мМ пирувата натрия (Gibco № 11360039), 100 мкг мл −1 Пенициллин, 100 мкг мл −1 Стрептомицин (Gibco # 15140122) и 100 мкг мл –1 Примоцин (InvivoGen # ant-pm-2)), при 32°C, % CO 2 , 3% O 2 кроме клеток мыши, которые инкубировали при 37 °C, 5% CO 2 . Для иммортализации фибробластов кожи ЯМР лентивирусная плазмида, несущая SV40LT (SV40LT-hRASg12v использовали для клеток мыши), была упакована с использованием HEK293FT в качестве упаковочных клеток. Собирали супернатант упаковочных клеток HEK293FT, содержащий вирусные частицы, упакованные SV40 LT.
Кожу кратко встряхивали каждые 30 минут, чтобы способствовать диссоциации клеток, и вручную проверяли диссоциацию клеток. После полной диссоциации клетки осаждали центрифугированием при 500 g в течение 5 минут и ресуспендировали в ЯМР среде для культивирования клеток (DMEM с высоким содержанием глюкозы (Gibco # 11965092) с добавлением 15% эмбриональной бычьей сыворотки (Gibco), заменимых аминокислот (Gibco # 11965092). 11140050), 1 мМ пирувата натрия (Gibco № 11360039), 100 мкг мл −1 Пенициллин, 100 мкг мл −1 Стрептомицин (Gibco # 15140122) и 100 мкг мл –1 Примоцин (InvivoGen # ant-pm-2)), при 32°C, % CO 2 , 3% O 2 кроме клеток мыши, которые инкубировали при 37 °C, 5% CO 2 . Для иммортализации фибробластов кожи ЯМР лентивирусная плазмида, несущая SV40LT (SV40LT-hRASg12v использовали для клеток мыши), была упакована с использованием HEK293FT в качестве упаковочных клеток. Собирали супернатант упаковочных клеток HEK293FT, содержащий вирусные частицы, упакованные SV40 LT. Когда первичные ЯМР-клетки кожи достигли 40% слияния, супернатант, содержащий вирусные частицы в упаковке SV40, добавляли к первичным ЯМР-клеткам кожи. Через 48 часов после добавления супернатанта клетки обрабатывали 2 мкг/мл пуромицина для уничтожения неинфицированных клеток. Стабильные иммортализованные клетки поддерживали в среде DMEM с добавлением 15% эмбриональной бычьей сыворотки, заменимых аминокислот, пирувата натрия, 100 ед. мл -1 пенициллина и 100 мг мл -1 стрептомицина.
Когда первичные ЯМР-клетки кожи достигли 40% слияния, супернатант, содержащий вирусные частицы в упаковке SV40, добавляли к первичным ЯМР-клеткам кожи. Через 48 часов после добавления супернатанта клетки обрабатывали 2 мкг/мл пуромицина для уничтожения неинфицированных клеток. Стабильные иммортализованные клетки поддерживали в среде DMEM с добавлением 15% эмбриональной бычьей сыворотки, заменимых аминокислот, пирувата натрия, 100 ед. мл -1 пенициллина и 100 мг мл -1 стрептомицина. Экстракция ГК из тканей методом ЯМР
 На следующий день образцы подвергали центрифугированию в холодном состоянии при 18 000×g в течение 10 минут. Затем гранулы промывали центрифугированием в четырех объемах 75% этанола. Супернатант удаляли, а осадки инкубировали при комнатной температуре в течение 20 минут для удаления остаточного этанола. Затем осадки ресуспендировали в 200 мкл автоклавированной дистиллированной воды. Добавляли 500 единиц бензоназной эндонуклеазы и затем инкубировали в течение ночи при 37 °C для удаления нуклеиновых кислот. На следующий день образцы осаждали в течение ночи одним объемом 100% этанола. На следующий день после холодного центрифугирования при 18000×g в течение 10 минут осадки ресуспендировали в 400 мкл автоклавированной дистиллированной воды.
На следующий день образцы подвергали центрифугированию в холодном состоянии при 18 000×g в течение 10 минут. Затем гранулы промывали центрифугированием в четырех объемах 75% этанола. Супернатант удаляли, а осадки инкубировали при комнатной температуре в течение 20 минут для удаления остаточного этанола. Затем осадки ресуспендировали в 200 мкл автоклавированной дистиллированной воды. Добавляли 500 единиц бензоназной эндонуклеазы и затем инкубировали в течение ночи при 37 °C для удаления нуклеиновых кислот. На следующий день образцы осаждали в течение ночи одним объемом 100% этанола. На следующий день после холодного центрифугирования при 18000×g в течение 10 минут осадки ресуспендировали в 400 мкл автоклавированной дистиллированной воды. Экстракция НА из кондиционированной среды
 После центрифугирования 2 мл кондиционированной среды инкубировали в течение ночи с 500 мкг протеиназы К при 50°С для удаления белков. На следующий день к образцам добавляли 2 объема 100% этанола для осаждения и инкубировали в течение ночи при -20 °С. На следующий день образцы подвергали центрифугированию в холодном состоянии при 18 000×g в течение 10 минут. После центрифугирования супернатант удаляли, а осадки инкубировали при комнатной температуре в течение 20 минут, чтобы дать возможность испариться остаткам этанола. После инкубации осадок ресуспендировали в 400 мкл автоклавированной дистиллированной воды.
После центрифугирования 2 мл кондиционированной среды инкубировали в течение ночи с 500 мкг протеиназы К при 50°С для удаления белков. На следующий день к образцам добавляли 2 объема 100% этанола для осаждения и инкубировали в течение ночи при -20 °С. На следующий день образцы подвергали центрифугированию в холодном состоянии при 18 000×g в течение 10 минут. После центрифугирования супернатант удаляли, а осадки инкубировали при комнатной температуре в течение 20 минут, чтобы дать возможность испариться остаткам этанола. После инкубации осадок ресуспендировали в 400 мкл автоклавированной дистиллированной воды. Экстракция ГК из кожи человека
 ). Кожу человека нарезали и взвешенное количество кожных тканей расщепляли в течение ночи при 50°C в буфере для расщепления (10 мМ Tris-Cl, 25 мМ EDTA, 100 мМ NaCl, 0,5% SDS и 0,1 мг мл 9).0009 -1
). Кожу человека нарезали и взвешенное количество кожных тканей расщепляли в течение ночи при 50°C в буфере для расщепления (10 мМ Tris-Cl, 25 мМ EDTA, 100 мМ NaCl, 0,5% SDS и 0,1 мг мл 9).0009 -1
Адсорбция HA на слюде
30 мкл очищенного NMRHA в воде наносили на свежерасщепленную мусковитную слюду и сушили на воздухе в течение 45 минут. Это метод, описанный Cowman и др. . для АСМ изображения HA 24 . Они отметили, что ГК слабо адсорбируется на слюде и не может быть исследована в жидких условиях из-за сродства ГК к раствору. Для исследования гидратированной ГК образцу давали высохнуть в течение 15 минут, чтобы видимая вода испарилась, но образец все еще оставался влажным.
Это метод, описанный Cowman и др. . для АСМ изображения HA 24 . Они отметили, что ГК слабо адсорбируется на слюде и не может быть исследована в жидких условиях из-за сродства ГК к раствору. Для исследования гидратированной ГК образцу давали высохнуть в течение 15 минут, чтобы видимая вода испарилась, но образец все еще оставался влажным.
Атомно-силовая микроскопия
Все изображения были получены на микроскопе Agilent 5500, оснащенном сканерами с обратной связью. Визуализация в контактном режиме использовалась для топографической визуализации с использованием кремниевых наконечников, имеющих номинальную постоянную силы в диапазоне 0,02–0,77 Н/м. Силы минимизировались при сканировании на уровне менее 1 нН. Частота сканирования составляла 0,5–1 кГц, и все изображения записывались с разрешением 512 пикселей. Измерения проводились в помещении со сверхчистой водой при комнатной температуре (~20 °C). Обработку и анализ изображений проводили с помощью программного обеспечения SPIP версии 6. 3.3 (Image Metrology, Lyngby, Дания).
3.3 (Image Metrology, Lyngby, Дания).
Наноиндентирование на основе атомно-силовой микроскопии
Использовались два различных типа геометрии индентирования кончика. Первые представляли собой конические наконечники с радиусом кривизны менее 15 нм. Они использовались для наблюдения за разворачивающимися событиями внутри суперспирали. Точное определение жесткости пружины было получено с использованием теоремы о равнораспределении, предложенной Hutter и Bechhoefer 34 . Неточности с точки зрения знания площади контакта между наконечником и образцом в модели Снеддона для конических наконечников можно компенсировать, используя большой сферический зонд, поэтому для определения значений модуля Юнга использовались сферические зонды. Сферические боросиликатные зонды диаметром 5 мкм (NovaScan Technologies) использовались для индентирования суперспиралей, как влажных, так и сухих. Пружинные константы определялись с использованием термического К-метода.
Доступность данных
Наборы данных, созданные в ходе и/или проанализированные в ходе текущего исследования, доступны у соответствующего автора по обоснованному запросу.
Ссылки
Гириш, К. С. и Кемпараю, К. Волшебный клей гиалуроновой кислоты и ее ластик гиалуронидаза: биологический обзор. Науки о жизни. 80 , 1921–1943 (2007).
КАС Статья Google ученый
Hargittai, I. & Hargittai, M. Молекулярная структура гиалуроновой кислоты: введение. Структура. хим. 19 , 697–717 (2008).
КАС Статья Google ученый
Рэнкин, К. С. и Франкель, Д. Гиалуронан при раке — от голого землекопа до терапии наночастицами. Soft Matter 12 , 3841–3848 (2016).
ОБЪЯВЛЕНИЕ КАС Статья Google ученый
Cheng, X.B., Sato, N., Kohi, S. & Yamaguchi, K. Прогностическое влияние гиалуронана и его регуляторов на аденокарциному протоков поджелудочной железы.
PLoS One 8 , 1–7 (2013).Google ученый
Аувинен, П. и др. . Повышенное содержание гиалуронана и CD44 стромальных клеток ассоциируется с HER2-позитивностью и неблагоприятным прогнозом при раке молочной железы человека. Междунар. Дж. Рак 132 , 531–539 (2013).
КАС Статья Google ученый
Сато Н., Кохи С., Хирата К. и Гоггинс М. Роль гиалуроновой кислоты в биологии и терапии рака поджелудочной железы: снова в центре внимания. Науки о раке. 107 , 569–575 (2016).
КАС Статья Google ученый
Тамми, Р. Х. и др. . Гиалуронан в опухолях человека: патобиологические и прогностические сообщения от клеточно-ассоциированного и стромального гиалуронана. Семин. Рак биол.
18 , 288–95 (2008).КАС Статья Google ученый
Риччарделли, К. и др. . Формирование перицеллюлярного матрикса, богатого гиалуроновой кислотой и версиканом, клетками рака предстательной железы способствует подвижности клеток. Дж. Биол. хим. 282 , 10814–10825 (2007 г.).
КАС Статья Google ученый
Qhattal, H.S.S. & Liu, X. Характеристика CD44-опосредованного захвата раковыми клетками и внутриклеточного распределения липосом с трансплантатом гиалуроновой кислоты. Мол. фарм. 8 , 1233–1246 (2011).
КАС Статья Google ученый
Кларк, Р. А., Алон, Р. и Спрингер, Т. А. CD44 и гиалуронан-зависимые вращающиеся взаимодействия лимфоцитов на строме миндалин. J. Cell Biol.
134 , 1075–1087 (1996).КАС Статья Google ученый
Richter, U., Wicklein, D., Geleff, S. & Schumacher, U. Взаимодействие между CD44 на опухолевых клетках и гиалуроновой кислотой в условиях физиологического потока: значение для образования метастазов. Гистохим. Клеточная биол. 137 , 687–695 (2012).
КАС Статья Google ученый
Senbanjo, L. T. & Chellaiah, M. A. CD44: Многофункциональный рецептор адгезии на клеточной поверхности является регулятором прогрессирования и метастазирования раковых клеток. Перед. Сотовый Дев. Биол . 5, (2017).
Cyphert, J.M., Trempus, C.S. & Garantziotis, S. Размер имеет значение: специфичность молекулярной массы эффектов гиалуроновой кислоты в клеточной биологии. Междунар. Дж. Селл Биол . 2015 г.
, https://doi.org/10.1155/2015/563818 (2015 г.).Fraser, J.R.E., Laurent, T.C. & Laurent, U.B.G. Гиалуронан: его природа, распределение, функции и оборот. Дж. Междунар. Мед. 242 , 27–33 (1997).
КАС Статья Google ученый
Руби, Дж. Г., Смит, М. и Баффенштейн, Р. Смертность голых землекопов противоречит законам Гомперца, поскольку не увеличивается с возрастом. Elife 7 , 1–18 (2018).
Артикул Google ученый
Шумахер, Л.-Н., Хассон, З. и Сент-Джон Смит, Э. Голый землекоп как животная модель в биомедицинских исследованиях: текущие перспективы. Анимация в открытом доступе. Физиол. 7 , 137–148 (2015).
Google ученый
Буффенштейн, Р. Незначительное старение у самого долгоживущего грызуна, голого землекопа: взгляды успешно стареющего вида.
Дж. Комп. Физиол. Б Биохим. Сист. Окружающая среда. Физиол. 178 , 439–445 (2008).Артикул Google ученый
Тянь, X. и др. . Гиалуронан с высокой молекулярной массой опосредует устойчивость голого землекопа к раку. Природа 499 , 346–349 (2013).
ОБЪЯВЛЕНИЕ КАС Статья Google ученый
Коуман М.К. и др. . Расширенные, релаксированные и конденсированные конформации гиалуронана, наблюдаемые с помощью атомно-силовой микроскопии. Биофиз. J. 88 , 590–602 (2005).
ОБЪЯВЛЕНИЕ КАС Статья Google ученый
Лю К., Ван М., Ан Дж., Торманн Э. и Дединайте А. Гиалуронан и фосфолипиды в граничной смазке. Soft Matter 8 , 10241–10244 (2012).
ОБЪЯВЛЕНИЕ КАС Статья Google ученый
Скотт, Дж. Э., Каммингс, К., Брасс, А. и Чен, Ю. Вторичные и третичные структуры гиалуронана в водном растворе, исследованные с помощью ротационной теневой электронной микроскопии и компьютерного моделирования. Биохим. J. 274 , 699–705 (1991).
КАС Статья Google ученый
Спаньоли, К. и др. . Конформации гиалуроновой кислоты на поверхности: эффект поверхностного заряда и гидрофобность. Углевод. Рез. 340 , 929–41 (2005).
КАС Статья Google ученый
Giannotti, M.I., Rinaudo, M. & Vancso, GJ. Силовая спектроскопия гиалуронана с помощью атомно-силовой микроскопии: от сетей с водородными связями к одноцепочечному поведению. Биомакромолекулы 8 , 2648–2652 (2007).
КАС Статья Google ученый
Луан Т., Ву Л., Чжан Х. и Ван Ю. Исследование природы межмолекулярных связей в криотропных слабых гелях гиалуронана. Углевод. Полим. 87 , 2076–2085 (2012).
КАС Статья Google ученый
Папаконстантину, Э.
, Рот, М. и Каракиулакис, Г. Гиалуроновая кислота: ключевая молекула в старении кожи. Дерматоэндокринол . 253–258, https://doi.org/10.4161/derm.21923 (2012).Рипеллино, Дж. А., Клингер, М. М., Марголис, Р. У. и Марголис, Р. К. Область связывания гиалуроновой кислоты как специфический зонд для локализации гиалуроновой кислоты в срезах тканей. J. Histochem. Цитохим. 33 , 1060–1066 (1985).
КАС Статья Google ученый
Lee, S.E., Jun, J.E., Choi, E.H., Ahn, S.K. & Lee, S.H. Стимуляция потери эпидермального градиента кальция увеличивает экспрессию гиалуроновой кислоты и CD44 в коже мыши. клин. Эксп. Дерматол. 35 , 650–657 (2010).
Артикул Google ученый
Crescenzi, V., Francescangeli, A., Renier, D. & Bellini, D. Новые сшитые и сульфатированные производные частично деацетилированного гиалуронана: синтез и предварительная характеристика.
Биополимеры 64 , 86–94 (2002).КАС Статья Google ученый
Ян, Ю. Л. и Кауфман, Л. Дж. Реология и конфокальная отражательная микроскопия как зонды механических свойств и структуры во время самосборки коллагена и коллагена/гиалуроновой кислоты. Биофиз. J. 96 , 1566–1585 (2009).
ОБЪЯВЛЕНИЕ КАС Статья Google ученый
Коуман, М.К., Ли, Х.-Г., Швертфегер, К.Л., Маккарти, Дж.Б. и Терли, Э.А. Содержание и размер гиалуроновой кислоты в биологических жидкостях и тканях. Перед. Иммунол. 6 , 261, https://doi.org/10.3389/fimmu.2015.00261 (2015).
КАС Статья пабмед ПабМед Центральный Google ученый
Каур, А. и др. . Ремоделирование коллагеновой матрицы в стареющей коже способствует метастазированию меланомы и влияет на подвижность иммунных клеток.
Открытие рака . https://doi.org/10.1158/2159-8290.CD-18-0193 (2019 г.).Артикул пабмед Google ученый
Хаттер, Дж. Л. и Бечхофер, Дж. Калибровка наконечников для атомно-силовой микроскопии. Rev.Sci. Инструм. 64 , 1868–1873 (1993).
ОБЪЯВЛЕНИЕ КАС Статья Google ученый
 PLoS One 8 , 1–7 (2013).
PLoS One 8 , 1–7 (2013). 18 , 288–95 (2008).
18 , 288–95 (2008). 134 , 1075–1087 (1996).
134 , 1075–1087 (1996). , https://doi.org/10.1155/2015/563818 (2015 г.).
, https://doi.org/10.1155/2015/563818 (2015 г.). Дж. Комп. Физиол. Б Биохим. Сист. Окружающая среда. Физиол. 178 , 439–445 (2008).
Дж. Комп. Физиол. Б Биохим. Сист. Окружающая среда. Физиол. 178 , 439–445 (2008). «>
«>Мураи, Т., Хоконохара, Х., Такаги, А. и Каваи, Т. Атомно-силовая микроскопия визуализации надмолекулярной организации гиалуроновой кислоты и ее рецептора CD44. IEEE Trans. Нанобиология 8 , 294–9 (2009).
Артикул Google ученый
 «>
«>Коуман М.К., Ли М. и Балаш Э.А. Атомно-силовая микроскопия гиалуроновой кислоты в режиме постукивания: расширенные и внутримолекулярные взаимодействующие цепи. Биофиз. Дж. 75 , 2030–2037 (1998).
ОБЪЯВЛЕНИЕ КАС Статья Google ученый
 , Рот, М. и Каракиулакис, Г. Гиалуроновая кислота: ключевая молекула в старении кожи. Дерматоэндокринол . 253–258, https://doi.org/10.4161/derm.21923 (2012).
, Рот, М. и Каракиулакис, Г. Гиалуроновая кислота: ключевая молекула в старении кожи. Дерматоэндокринол . 253–258, https://doi.org/10.4161/derm.21923 (2012). Биополимеры 64 , 86–94 (2002).
Биополимеры 64 , 86–94 (2002). Открытие рака . https://doi.org/10.1158/2159-8290.CD-18-0193 (2019 г.).
Открытие рака . https://doi.org/10.1158/2159-8290.CD-18-0193 (2019 г.).Ссылки на скачивание
Благодарности
Эта работа была поддержана наградой за многодисциплинарный проект Cancer Research UK/RCUK (C56829/A22053) для K.R., E.S. и Д.Ф. и награда за учреждение карьеры в Великобритании по исследованию рака (C47525 / A17348) для W.T.K. FH и SC были поддержаны стипендиями Gates Cambridge Trust.
Информация об авторе
Авторы и представители
Факультет фармакологии Кембриджского университета, Tennis Court Road, Cambridge, CB2 1PD, UK
Явуз Кулабероглу, Фазал Хади, Сампурна Чакрабарти, Валид Т.
Халед и Эван Сент-Джон Смит1
Инженерная школа Университета Ньюкасла, Мерц Корт, Ньюкасл-апон-Тайн, NE1 7RU, Великобритания
Бхарат Бхушан и Даниэль Франкель
Северный институт исследований рака, Медицинская школа, Университет Ньюкасла, Ньюкасл-апон-Тайн, NE2 4HH, Великобритания
Kenneth S. Rankin
 Халед и Эван Сент-Джон Смит
Халед и Эван Сент-Джон СмитАвторы
- Явуз Кулабероглу
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Bharat Bhushan
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Fazal Hadi
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Академия
- Сампурна Чакрабарти
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Валид Т. Халед
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Kenneth S. Rankin
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Академия
- Ewan St. John Smith
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Daniel Frankel
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
 Халед
ХаледContributions
E.S., K.R. и Д.Ф. задумал исследование, проанализировал данные и написал рукопись. Ю.К., Б.Б. и Д.Ф. провели эксперименты по извлечению и характеристике H.A.S.C. провели иммуногистохимические эксперименты. В.К. и FH отвечали за создание клеточных линий.
Авторы переписки
Переписка с
Кеннет С. Рэнкин, Юэн Сент-Джон Смит или Дэниел Франкель.
Заявление об этике
Конкурирующие интересы
Авторы не заявляют об отсутствии конкурирующих интересов.
Дополнительная информация
Примечание издателя: Springer Nature остается нейтральной в отношении юрисдикционных претензий в опубликованных картах и институциональной принадлежности.
Права и разрешения
Открытый доступ Эта статья находится под лицензией Creative Commons Attribution 4.0 International License, которая разрешает использование, совместное использование, адаптацию, распространение и воспроизведение на любом носителе или в любом формате при условии, что вы укажете автора(ов) оригинала и источник, предоставьте ссылку на лицензию Creative Commons и укажите, были ли внесены изменения. Изображения или другие сторонние материалы в этой статье включены в лицензию Creative Commons для статьи, если иное не указано в кредитной строке материала. Если материал не включен в лицензию Creative Commons статьи, а ваше предполагаемое использование не разрешено законом или превышает разрешенное использование, вам необходимо получить разрешение непосредственно от правообладателя. Чтобы просмотреть копию этой лицензии, посетите http://creativecommons.org/licenses/by/4.0/.
Чтобы просмотреть копию этой лицензии, посетите http://creativecommons.org/licenses/by/4.0/.
Перепечатка и разрешения
Об этой статье
Дополнительная литература
Содержание и размер гиалуроновой кислоты в тканях и плазме голого землекопа
- Дельфин дель Мармоль
- Сюзанна Хольце
- Бруно Фламион
Научные отчеты (2021)
Нетрадиционные системы в исследованиях старения: обновление
- Юстина Микула-Петрасик
- Мартина Пакула
- Кшиштоф Ксенжек
Клеточные и молекулярные науки о жизни (2021)
Трансформация клеток голого землекопа
- Фазал Хади
- Явуз Кулабероглу
- Валид Т. Халед
Природа (2020)
 Халед
ХаледКомментарии
Отправляя комментарий, вы соглашаетесь соблюдать наши Условия и Правила сообщества. Если вы обнаружите что-то оскорбительное или не соответствующее нашим условиям или правилам, отметьте это как неприемлемое.
Гиалуронан с очень высокой молекулярной массой голого землекопа проявляет превосходные цитопротекторные свойства ), по сравнению с другими видами млекопитающих. Однако неясно, важна ли исключительная длина полимера vHMM-HA для долголетия. Здесь мы показываем, что vHMM-HA (> 6,1 МДа) обладает превосходными цитопротекторными свойствами по сравнению с более коротким HMM-HA. Он защищает не только клетки ЯМР, но также клетки мыши и человека от индуцированной стрессом остановки клеточного цикла и гибели клеток в зависимости от длины полимера.
 Цитопротекторный эффект зависит от основного HA-рецептора, CD44. Мы обнаружили, что vHMM-HA подавляет белок-белковые взаимодействия CD44, тогда как HMM-HA стимулирует их. В результате vHMM-HA и HMM-HA вызывают противоположные эффекты на экспрессию CD44-зависимых генов, которые связаны с путем p53. Одновременно vHMM-HA частично ослабляет р53 и защищает клетки от стресса р53-зависимым образом. Наши результаты указывают на участие vHMM-HA в механизмах замедления старения и предполагают потенциальное применение vHMM-HA для повышения устойчивости клеток к стрессу.
Цитопротекторный эффект зависит от основного HA-рецептора, CD44. Мы обнаружили, что vHMM-HA подавляет белок-белковые взаимодействия CD44, тогда как HMM-HA стимулирует их. В результате vHMM-HA и HMM-HA вызывают противоположные эффекты на экспрессию CD44-зависимых генов, которые связаны с путем p53. Одновременно vHMM-HA частично ослабляет р53 и защищает клетки от стресса р53-зависимым образом. Наши результаты указывают на участие vHMM-HA в механизмах замедления старения и предполагают потенциальное применение vHMM-HA для повышения устойчивости клеток к стрессу.Введение
Самый долгоживущий грызун, голый землекоп (ЯМР) ( Heterocephalus glaber ), имеет максимальную продолжительность жизни более 30 лет, что в пять раз больше, чем предсказывает масса тела 1 . ЯМР не показывает повышения уровня смертности в течение по крайней мере 18 лет 2 и, кажется, защищен от возрастных ухудшений, таких как снижение метаболизма, диабет и остеопороз 3,4 Более того, репродуктивная функция женского ЯМР даже увеличивается с возрастом до 20 лет 9 лет0009 4 . Эти особенности указывают на то, что ЯМР развил эффективные механизмы против старения. Однако, хотя ЯМР все больше ценят как модель для исследования старения, остается в значительной степени неизвестным, как они сопротивляются процессам старения. Интересно, что хотя базальные уровни окислительного стресса выше в клетках ЯМР, чем в клетках мыши 5,6 , связанное с возрастом увеличение окислительного повреждения намного меньше в ЯМР 7 . Кроме того, белки ЯМР лучше сохраняют свою структуру и функции в условиях окислительного стресса 6,7 , что хотя бы частично можно объяснить высокой активностью протеасом и аутофагии в клетках ЯМР 8,9 . Эти наблюдения показывают, что управление клеточным стрессом, а не подавление самого повреждения, имеет решающее значение для долговечности ЯМР. В соответствии с этой идеей мы недавно обнаружили, что одна и та же доза облучения вызывает аналогичные повреждения ДНК, но меньшее клеточное старение в клетках ЯМР, чем в клетках мыши 10 .
Эти особенности указывают на то, что ЯМР развил эффективные механизмы против старения. Однако, хотя ЯМР все больше ценят как модель для исследования старения, остается в значительной степени неизвестным, как они сопротивляются процессам старения. Интересно, что хотя базальные уровни окислительного стресса выше в клетках ЯМР, чем в клетках мыши 5,6 , связанное с возрастом увеличение окислительного повреждения намного меньше в ЯМР 7 . Кроме того, белки ЯМР лучше сохраняют свою структуру и функции в условиях окислительного стресса 6,7 , что хотя бы частично можно объяснить высокой активностью протеасом и аутофагии в клетках ЯМР 8,9 . Эти наблюдения показывают, что управление клеточным стрессом, а не подавление самого повреждения, имеет решающее значение для долговечности ЯМР. В соответствии с этой идеей мы недавно обнаружили, что одна и та же доза облучения вызывает аналогичные повреждения ДНК, но меньшее клеточное старение в клетках ЯМР, чем в клетках мыши 10 .
Помимо незначительного старения организма, ЯМР показывает чрезвычайно высокую устойчивость к раку. Клетки ЯМР экспрессируют уникальную изоформу INK4, названную pALT 9.0009 INK4a/b , который активируется при различных стрессах и обладает более сильной способностью индуцировать остановку клеточного цикла, чем p15 INK4b или p16 INK4a 11 . Также сообщалось, что потеря ARF запускает клеточное старение, не влияя на уровни экспрессии p16 INK4A , p21 Cip1/Waf1 и p27 Kip1 в клетках ЯМР 12 . Эти видоспецифические функции генов-супрессоров опухолей могут способствовать устойчивости к раку в ЯМР. Другим важным противораковым механизмом, специфичным для ЯМР, является раннее контактное ингибирование (ECI) 9.0009 13,14,15 . Культивируемые ЯМР-фибробласты гиперчувствительны к контактному ингибированию и прекращают пролиферацию при относительно низкой плотности клеток зависимым от гиалуроновой кислоты (HA) образом. HA, линейный полисахарид и основной компонент внеклеточного матрикса, играет роль в поддержании структуры ткани и регуляции клеточных сигнальных путей в зависимости от длины его полимера 15,16 . Динамическая регуляция количества и длины полимера ГК участвует в различных биологических процессах, включая пролиферацию клеток, миграцию клеток и воспаление. В здоровых тканях большая часть ГК имеет высокую молекулярную массу ( M > 1 МДа) (HMM-HA). При патологических состояниях происходит значительная фрагментация ГК с образованием низкомолекулярной ГК ( M < 250 кДа) 15,17 . ЯМР производит гиалуронан с очень высокой молекулярной массой (vHMM-HA), намного дольше, чем HA у других видов млекопитающих 14 . Однако до сих пор неясно, отличается ли функционально ГА исключительно большой длины полимера от обычного ГММ-ГА. Одним из механизмов, лежащих в основе зависимой от длины полимера передачи сигнала HA, является дифференциальная кластеризация рецепторов HA, запускаемая их поливалентным взаимодействием с HA 9.
HA, линейный полисахарид и основной компонент внеклеточного матрикса, играет роль в поддержании структуры ткани и регуляции клеточных сигнальных путей в зависимости от длины его полимера 15,16 . Динамическая регуляция количества и длины полимера ГК участвует в различных биологических процессах, включая пролиферацию клеток, миграцию клеток и воспаление. В здоровых тканях большая часть ГК имеет высокую молекулярную массу ( M > 1 МДа) (HMM-HA). При патологических состояниях происходит значительная фрагментация ГК с образованием низкомолекулярной ГК ( M < 250 кДа) 15,17 . ЯМР производит гиалуронан с очень высокой молекулярной массой (vHMM-HA), намного дольше, чем HA у других видов млекопитающих 14 . Однако до сих пор неясно, отличается ли функционально ГА исключительно большой длины полимера от обычного ГММ-ГА. Одним из механизмов, лежащих в основе зависимой от длины полимера передачи сигнала HA, является дифференциальная кластеризация рецепторов HA, запускаемая их поливалентным взаимодействием с HA 9. 0009 16,18 Многочисленные исследования показали, что низко-, но не HMM-HA способствует воспалительным реакциям 19 . С другой стороны, HMM-HA повышает устойчивость клеток к стрессу, особенно к окислительному стрессу 20,21,22,23 , хотя лежащие в его основе механизмы недостаточно изучены. Одно предостережение здесь заключается в том, что концентрации ГК, использованные в этих исследованиях, были относительно высокими; более 100 мкг/мл, во многих случаях 1–2 мкг/мл, тогда как в большинстве органов, кроме дермы и хрящей, содержание ГК составляет 1–100 мкг ГК/г влажной ткани 17,24 .
0009 16,18 Многочисленные исследования показали, что низко-, но не HMM-HA способствует воспалительным реакциям 19 . С другой стороны, HMM-HA повышает устойчивость клеток к стрессу, особенно к окислительному стрессу 20,21,22,23 , хотя лежащие в его основе механизмы недостаточно изучены. Одно предостережение здесь заключается в том, что концентрации ГК, использованные в этих исследованиях, были относительно высокими; более 100 мкг/мл, во многих случаях 1–2 мкг/мл, тогда как в большинстве органов, кроме дермы и хрящей, содержание ГК составляет 1–100 мкг ГК/г влажной ткани 17,24 .
Наблюдения за тем, что ГК проявляет цитозащитный эффект, зависящий от длины полимера, и что долгоживущий ЯМР продуцирует vHMM-HA, приводят к гипотезе о том, что дополнительная длина полимера ЯМР-ГК обеспечивает превосходную цитозащиту, которая может способствовать долговечности ЯМР. Здесь мы показываем, что vHMM-HA обладает превосходными цитопротекторными свойствами по сравнению с более коротким HMM-HA. Механически vHMM-HA и HMM-HA оказывают противоположное влияние на белок-белковые взаимодействия CD44 (PPI) и CD44-зависимую экспрессию генов. В результате vHMM-HA частично ослабляет путь р53 и защищает клетки от индуцированной стрессом остановки клеточного цикла и гибели клеток.
Механически vHMM-HA и HMM-HA оказывают противоположное влияние на белок-белковые взаимодействия CD44 (PPI) и CD44-зависимую экспрессию генов. В результате vHMM-HA частично ослабляет путь р53 и защищает клетки от индуцированной стрессом остановки клеточного цикла и гибели клеток.
Результаты
ЯМР-ГК защищает клетки от окислительного стресса посредством CD44
Чтобы изучить потенциальные цитозащитные эффекты ЯМР-ГК, мы сначала проверили, защищает ли ауто/паракринное действие ГК, секретируемой кожными фибробластами ЯМР (NSF), от Гибель клеток, индуцированная tert -бутилгидропероксидазой (tBHP). tBHP является широко используемым окислительным стрессором, который продуцирует пероксильные/алкоксильные радикалы или истощает запасы GSH посредством метаболизма. Как и ожидалось, фермент, разлагающий НА, Streptomyces гиалуронидаза (HAase) снижала жизнеспособность NSF через 2 дня лечения tBHP (дополнительная рис. 1A). Кроме того, кондиционированная среда (CM) NSF, но не среда фибробластов кожи мыши (MSF), подавляла гибель клеток хорошо охарактеризованных первичных фибробластов легких человека (клетки IMR90) после 2 дней обработки tBHP в HA- зависимым образом (дополнительный рис. 1B).
1B).
ГК может оказывать цитопротекторное действие, непосредственно удаляя АФК во внеклеточном пространстве или запуская внутриклеточные цитопротекторные сигнальные пути. Чтобы проверить, защищает ли NMR-HA клетки за счет повышения устойчивости клеток к стрессу, а не за счет удаления АФК, мы предварительно инкубировали IMR9.0 с 20 мкг/мл (физиологическая концентрация во многих тканях) очищенного NSF-HA или эквивалентного объема PBS в течение 6 часов, а затем удаляли среду, содержащую HA или PBS, и обрабатывали клетки высокой дозой tBHP в течение 1 часа. Таким образом, ГК не присутствовала во время обработки tBHP, что устраняло ее прямое действие по очистке от АФК. Как показано на рис. 1а, 6-часовой предварительной инкубации с NSF-HA было достаточно, чтобы подавить гибель клеток, вызванную tBHP. Ежедневное повторение этих обработок с использованием низких доз tBHP вместо высоких доз приводило к зависимому от NSF-HA восстановлению пролиферации клеток (рис. 1b). Без обработки tBHP NSF-HA не стимулировал пролиферацию клеток и не индуцировал ECI-подобную остановку клеточного цикла в IMR9. 0 клеток, что указывает на то, что NSF-HA сам по себе не влияет на клеточный цикл (дополнительный рисунок 1C). Предварительная инкубация NSF-HA также уменьшала количество очагов повреждения ДНК после повторного лечения низкими дозами tBHP (рис. 1c; тест Уилкоксона p = 6,8 × 10 -6 ). Затем мы исследовали участие двух основных рецепторов HA, CD44 и RHAMM, в цитопротекторном эффекте NSF-HA. Нокдаун RHAMM не блокировал цитопротекторный эффект NSF-HA, тогда как миРНК CD44, а также нейтрализующее антитело против CD44 отменяли его эффект (рис. 1d, e и дополнительная рис. 2). Эти результаты показывают, что передача сигналов HA-CD44 отвечает за цитопротекцию с помощью NSF-HA.
0 клеток, что указывает на то, что NSF-HA сам по себе не влияет на клеточный цикл (дополнительный рисунок 1C). Предварительная инкубация NSF-HA также уменьшала количество очагов повреждения ДНК после повторного лечения низкими дозами tBHP (рис. 1c; тест Уилкоксона p = 6,8 × 10 -6 ). Затем мы исследовали участие двух основных рецепторов HA, CD44 и RHAMM, в цитопротекторном эффекте NSF-HA. Нокдаун RHAMM не блокировал цитопротекторный эффект NSF-HA, тогда как миРНК CD44, а также нейтрализующее антитело против CD44 отменяли его эффект (рис. 1d, e и дополнительная рис. 2). Эти результаты показывают, что передача сигналов HA-CD44 отвечает за цитопротекцию с помощью NSF-HA.
a NSF-HA подавляет вызванную окислительным стрессом гибель клеток IMR90. Процент живых клеток измеряли путем окрашивания аннексином-V/PI через 1 день после 1 часа обработки 3 мМ tBHP ( n = 3). Клетки предварительно инкубировали в течение 6 ч с 20 мкг/мл NSF-HA или PBS перед обработкой tBHP. b NSF-HA подавляет индуцированную окислительным стрессом остановку роста в клетках IMR90 ( n = 4). Клетки предварительно инкубировали в течение 6 часов с 20 мкг/мл NSF-HA или PBS, затем удаляли HA и клетки подвергали воздействию 200 мкМ tBHP в течение 1 часа. Эти процедуры повторяли ежедневно в течение первых 4 дней. 9Клетки 0065c IMR90 окрашивали антителами γh3AX и 53BP1 на 8-й день эксперимента, представленного в (b ). Гистограммы показывают распределение числа колокализованных очагов γh3AX/53BP1 на клетку. Для каждой группы образцов эксперимент повторяли ( n = 2) и всего подсчитывали более 100 клеток. Масштабные линейки, 20 мкм. d Антитело к CD44 блокирует цитопротекторный эффект NSF-HA на гибель клеток, вызванную окислительным стрессом, в клетках IMR90. Процент живых клеток измеряли с помощью окрашивания аннексином-V/PI через 1 день после 1 часа обработки 3 мМ tBHP (9).
Клетки предварительно инкубировали в течение 6 ч с 20 мкг/мл NSF-HA или PBS перед обработкой tBHP. b NSF-HA подавляет индуцированную окислительным стрессом остановку роста в клетках IMR90 ( n = 4). Клетки предварительно инкубировали в течение 6 часов с 20 мкг/мл NSF-HA или PBS, затем удаляли HA и клетки подвергали воздействию 200 мкМ tBHP в течение 1 часа. Эти процедуры повторяли ежедневно в течение первых 4 дней. 9Клетки 0065c IMR90 окрашивали антителами γh3AX и 53BP1 на 8-й день эксперимента, представленного в (b ). Гистограммы показывают распределение числа колокализованных очагов γh3AX/53BP1 на клетку. Для каждой группы образцов эксперимент повторяли ( n = 2) и всего подсчитывали более 100 клеток. Масштабные линейки, 20 мкм. d Антитело к CD44 блокирует цитопротекторный эффект NSF-HA на гибель клеток, вызванную окислительным стрессом, в клетках IMR90. Процент живых клеток измеряли с помощью окрашивания аннексином-V/PI через 1 день после 1 часа обработки 3 мМ tBHP (9). 0023 n = 3). Клетки предварительно инкубировали в течение 6 ч с 20 мкг/мл NSF-HA или PBS с блокирующим антителом CD44 или без него (10 нг/мл) перед обработкой tBHP. Антитело e CD44 блокирует цитопротективное действие NSF-HA на индуцированную окислительным стрессом остановку роста в клетках IMR90 ( n = 4). Клетки предварительно инкубировали в течение 6 часов с 20 мкг/мл NSF-HA или PBS с блокирующим антителом против CD44 или без него (10 нг/мл), затем HA удаляли и клетки подвергали воздействию 200 мкМ tBHP в течение 1 часа в день 0 и день 1. Масштабные линейки, 20 мкм. Столбики погрешностей представлены в виде средних значений ± SD. * p < 0,05 [двухсторонний t -критерий для ( a , b ) и однофакторный ANOVA с post-hoc Двусторонний критерий Даннета для ( 6 5 3 , d 9006 ].
0023 n = 3). Клетки предварительно инкубировали в течение 6 ч с 20 мкг/мл NSF-HA или PBS с блокирующим антителом CD44 или без него (10 нг/мл) перед обработкой tBHP. Антитело e CD44 блокирует цитопротективное действие NSF-HA на индуцированную окислительным стрессом остановку роста в клетках IMR90 ( n = 4). Клетки предварительно инкубировали в течение 6 часов с 20 мкг/мл NSF-HA или PBS с блокирующим антителом против CD44 или без него (10 нг/мл), затем HA удаляли и клетки подвергали воздействию 200 мкМ tBHP в течение 1 часа в день 0 и день 1. Масштабные линейки, 20 мкм. Столбики погрешностей представлены в виде средних значений ± SD. * p < 0,05 [двухсторонний t -критерий для ( a , b ) и однофакторный ANOVA с post-hoc Двусторонний критерий Даннета для ( 6 5 3 , d 9006 ].
Изображение в натуральную величину
vHMM-HA обладает превосходными цитозащитными свойствами
Чтобы оценить, способствует ли исключительная длина полимера NSF-HA его цитозащитному эффекту, мы предварительно инкубировали клетки IMR90 с NSF-HA или таким же количеством (20 мкг /мл) MSF-HA за 6 часов до лечения tBHP. Большая часть NSF-HA представляла собой vHMM-HA с молекулярной массой выше 6,1 МДа, тогда как вся MSF-HA была меньше 6,1 МДа (рис. 2а), как сообщалось ранее 14 . В отличие от NSF-HA, MSF-HA не повышал устойчивость к окислительному стрессу в клетках IMR90 (рис. 2b, c), хотя средний молекулярный размер MSF-HA по-прежнему относится к классу HMM-HA. Чтобы исключить возможность того, что это различие связано с примесями в двух препаратах ГК, мы затем сравнили влияние интактного и частично фрагментированного NSF-HA (fNSF-HA) на устойчивость клеток к стрессу. Для частичной фрагментации NSF-HA инкубировали с низкой концентрацией HAase в течение короткого периода времени, и реакцию останавливали путем инактивации фермента нагреванием. Для контроля NSF-HA нагревали после смешивания с термоинактивированной HAase. Следовательно, контрольные NSF-HA (cNSF-HA) и fNSF-HA должны быть точно идентичными, за исключением длины полимера HA. Хотя большая часть fNSF-HA сохранила молекулярную массу выше 1 МДа, она больше не защищала IMR9.
Большая часть NSF-HA представляла собой vHMM-HA с молекулярной массой выше 6,1 МДа, тогда как вся MSF-HA была меньше 6,1 МДа (рис. 2а), как сообщалось ранее 14 . В отличие от NSF-HA, MSF-HA не повышал устойчивость к окислительному стрессу в клетках IMR90 (рис. 2b, c), хотя средний молекулярный размер MSF-HA по-прежнему относится к классу HMM-HA. Чтобы исключить возможность того, что это различие связано с примесями в двух препаратах ГК, мы затем сравнили влияние интактного и частично фрагментированного NSF-HA (fNSF-HA) на устойчивость клеток к стрессу. Для частичной фрагментации NSF-HA инкубировали с низкой концентрацией HAase в течение короткого периода времени, и реакцию останавливали путем инактивации фермента нагреванием. Для контроля NSF-HA нагревали после смешивания с термоинактивированной HAase. Следовательно, контрольные NSF-HA (cNSF-HA) и fNSF-HA должны быть точно идентичными, за исключением длины полимера HA. Хотя большая часть fNSF-HA сохранила молекулярную массу выше 1 МДа, она больше не защищала IMR9. 0 клеток от стресса, индуцированного tBHP (рис. 2d – f). Обратите внимание, что распределение размеров молекул cNSF- и fNSF-HA не изменилось во время инкубации с клетками IMR90, что указывает на то, что отсутствие цитозащитного эффекта fNSF-HA не связано с деградацией HMM-HA во время эксперимента (дополнительная рис. 3А). Кроме того, cNSF-HA, но не fNSF-HA, защищал от остановки клеточного цикла, вызванной доксорубицином (DXR) и облучением, в клетках IMR90 (дополнительная рис. 3B, C). MSF также был защищен NSF-HA в зависимости от длины полимера (дополнительный рисунок 3D, E). Наконец, мы сравнили цитопротекторный эффект извлеченного из геля vHMM-HA (>6,1 MДа) и синтетического гиалуронана (Select-HA 9).0009 TM ) с однородной молекулярной массой 1 МДа. Экстрагированный из геля vHMM-HA защищал клетки IMR90 от окислительного стресса, но 1 МДа HA не защищал даже при пятикратно более высокой концентрации (дополнительная рис. 3F-I). Эти результаты показывают, что vHMM-HA обладает лучшими цитопротекторными свойствами по сравнению с более коротким HMM-HA.
0 клеток от стресса, индуцированного tBHP (рис. 2d – f). Обратите внимание, что распределение размеров молекул cNSF- и fNSF-HA не изменилось во время инкубации с клетками IMR90, что указывает на то, что отсутствие цитозащитного эффекта fNSF-HA не связано с деградацией HMM-HA во время эксперимента (дополнительная рис. 3А). Кроме того, cNSF-HA, но не fNSF-HA, защищал от остановки клеточного цикла, вызванной доксорубицином (DXR) и облучением, в клетках IMR90 (дополнительная рис. 3B, C). MSF также был защищен NSF-HA в зависимости от длины полимера (дополнительный рисунок 3D, E). Наконец, мы сравнили цитопротекторный эффект извлеченного из геля vHMM-HA (>6,1 MДа) и синтетического гиалуронана (Select-HA 9).0009 TM ) с однородной молекулярной массой 1 МДа. Экстрагированный из геля vHMM-HA защищал клетки IMR90 от окислительного стресса, но 1 МДа HA не защищал даже при пятикратно более высокой концентрации (дополнительная рис. 3F-I). Эти результаты показывают, что vHMM-HA обладает лучшими цитопротекторными свойствами по сравнению с более коротким HMM-HA.
a Изображение гель-электрофореза в импульсном поле NSF-HA и MSF-HA. Эксперимент был повторен один раз с аналогичным результатом. b NSF-HA, но не MSF-HA, подавляет гибель клеток, вызванную окислительным стрессом, в клетках IMR90. Процент живых клеток измеряли путем окрашивания аннексином-V/PI через 1 день после 1 часа обработки 3 мМ tBHP ( n = 3). Клетки предварительно инкубировали в течение 6 ч с 20 мкг/мл NSF-HA, MSF-HA или PBS перед обработкой tBHP. c NSF-HA, но не MSF-HA, подавляет индуцированную окислительным стрессом остановку роста в клетках IMR90 ( n = 4). Клетки предварительно инкубировали в течение 6 ч с 20 мкг/мл NSF-HA, MSF-HA или PBS, затем ГК удаляли и клетки подвергали воздействию 200 мкМ tBHP в течение 1 ч в день 0 и день 1. d Изображение гель-электрофореза в импульсном поле cNSF-HA и fNSF-HA. Эксперимент был повторен один раз с аналогичным результатом. e cNSF-HA, но не fNSF-HA, подавляет вызванную окислительным стрессом гибель клеток IMR90. Процент живых клеток измеряли путем окрашивания аннексином-V/PI через 1 день после 1 часа обработки 3 мМ tBHP ( n = 3). Клетки предварительно инкубировали в течение 6 часов с 20 мкг/мл cNSF-HA, fNSF-HA или PBS перед обработкой tBHP. f cNSF-HA, но не fNSF-HA, подавляет индуцированную окислительным стрессом остановку роста IMR90 ячеек ( n = 4). Клетки предварительно инкубировали в течение 6 часов с 20 мкг/мл cNSF-HA, fNSF-HA или PBS, затем удаляли HA и подвергали клетки воздействию 200 мкМ tBHP в течение 1 часа в день 0 и день 1. г Тепловая карта показывает нормализованное по CD44 содержание 99 ко-иммунопреципитированных белков CD44, которые по-разному связаны с CD44 в клетках IMR90 со сверхэкспрессией CD44. Клетки инкубировали в течение 6 часов с 20 мкг/мл cNSF-HA, fNSF-HA или PBS. Эксперимент проводился в трехкратной повторности. ( ч ) Вестерн-блоты, показывающие обилие коиммунопреципитированного CD44-FLAG CD44-YFP в клетках IMR90, сверхэкспрессирующих CD44-FLAG и CD44-YFP ( n = 9).
e cNSF-HA, но не fNSF-HA, подавляет вызванную окислительным стрессом гибель клеток IMR90. Процент живых клеток измеряли путем окрашивания аннексином-V/PI через 1 день после 1 часа обработки 3 мМ tBHP ( n = 3). Клетки предварительно инкубировали в течение 6 часов с 20 мкг/мл cNSF-HA, fNSF-HA или PBS перед обработкой tBHP. f cNSF-HA, но не fNSF-HA, подавляет индуцированную окислительным стрессом остановку роста IMR90 ячеек ( n = 4). Клетки предварительно инкубировали в течение 6 часов с 20 мкг/мл cNSF-HA, fNSF-HA или PBS, затем удаляли HA и подвергали клетки воздействию 200 мкМ tBHP в течение 1 часа в день 0 и день 1. г Тепловая карта показывает нормализованное по CD44 содержание 99 ко-иммунопреципитированных белков CD44, которые по-разному связаны с CD44 в клетках IMR90 со сверхэкспрессией CD44. Клетки инкубировали в течение 6 часов с 20 мкг/мл cNSF-HA, fNSF-HA или PBS. Эксперимент проводился в трехкратной повторности. ( ч ) Вестерн-блоты, показывающие обилие коиммунопреципитированного CD44-FLAG CD44-YFP в клетках IMR90, сверхэкспрессирующих CD44-FLAG и CD44-YFP ( n = 9). Клетки инкубировали в течение 6 часов с 20 мкг/мл cNSF-HA, fNSF-HA или PBS. i Количественная оценка Вестерн-блоттинга показана в ( h ) на изображении J. Столбики погрешностей представлены как среднее ± ± стандартное отклонение. * p < 0,05 (односторонний ANOVA с апостериорным двусторонним критерием Даннета).
Клетки инкубировали в течение 6 часов с 20 мкг/мл cNSF-HA, fNSF-HA или PBS. i Количественная оценка Вестерн-блоттинга показана в ( h ) на изображении J. Столбики погрешностей представлены как среднее ± ± стандартное отклонение. * p < 0,05 (односторонний ANOVA с апостериорным двусторонним критерием Даннета).
Полноразмерное изображение
vHMM- и HMM-HA оказывают противоположное влияние на интерактом CD44
Тот факт, что цитопротекторный эффект NSF-HA зависит от CD44 и его исключительной длины полимера, предполагает, что vHMM-HA и HMM-HA регулируют сигналы CD44 по-разному. Чтобы проверить это мнение, мы проанализировали влияние ГК на белок-белковые взаимодействия CD44. Мы сверхэкспрессировали стандартную форму CD44 в клетках IMR90 (дополнительная фигура 4A) и провели анализ коиммунопреципитации CD44 через 6 ч после инкубации с cNSF-HA, fNSF-HA или PBS. Большинство эндогенных молекул CD44 имели стандартную форму в IMR9.0, MSF 10 и NSF (дополнительный рисунок 4B, C). CD44 и его интеракторы перекрестно связывали с DSP и DTME перед сбором клеток. Затем иммунопреципитированные белки подвергали количественной масс-спектрометрии с тандемной масс-меткой (ТМТ). Это выявило 290 ко-иммунопреципитированных белков с обогащением как минимум в два раза по сравнению с контролем IgG (дополнительный набор данных 1). Из них 109 белков (38%) уже описаны как белки, взаимодействующие с CD44 25 , что указывает на успех коиммунопреципитации. Мы сравнили количество ко-иммунопреципитированных белков CD44 после нормализации до уровней CD44 и определили 99 белков должны по-разному ассоциироваться с CD44 в клетках IMR90, инкубированных с cNSF-HA и fNSF-HA ( p < 0,05). Поразительно, но эффективность коиммунопреципитации всех этих белков была выше в клетках, инкубированных с fNSF-HA, чем в клетках, инкубированных с cNSF-HA (дополнительный набор данных 1). Более того, по сравнению с PBS, cNSF-HA снижал эффективность коиммунопреципитации большинства этих белков, тогда как fNSF-HA в основном усиливал ее (рис.
CD44 и его интеракторы перекрестно связывали с DSP и DTME перед сбором клеток. Затем иммунопреципитированные белки подвергали количественной масс-спектрометрии с тандемной масс-меткой (ТМТ). Это выявило 290 ко-иммунопреципитированных белков с обогащением как минимум в два раза по сравнению с контролем IgG (дополнительный набор данных 1). Из них 109 белков (38%) уже описаны как белки, взаимодействующие с CD44 25 , что указывает на успех коиммунопреципитации. Мы сравнили количество ко-иммунопреципитированных белков CD44 после нормализации до уровней CD44 и определили 99 белков должны по-разному ассоциироваться с CD44 в клетках IMR90, инкубированных с cNSF-HA и fNSF-HA ( p < 0,05). Поразительно, но эффективность коиммунопреципитации всех этих белков была выше в клетках, инкубированных с fNSF-HA, чем в клетках, инкубированных с cNSF-HA (дополнительный набор данных 1). Более того, по сравнению с PBS, cNSF-HA снижал эффективность коиммунопреципитации большинства этих белков, тогда как fNSF-HA в основном усиливал ее (рис. 2g). Эти результаты показывают, что белок-белковые взаимодействия CD44 стимулируются HMM-HA, но подавляются vHMM-HA. Затем, чтобы прояснить влияние vHMM-HA на белковое взаимодействие CD44-CD44, мы одновременно сверхэкспрессировали CD44-FLAG и CD44-YFP в IMR9.0 клеток и исследовали их взаимодействие с помощью анализа коиммунопреципитации FLAG. В соответствии с нашими данными масс-спектрометрии, cNSF-HA уменьшал количество коиммунопреципитированного CD44-YFP, тогда как fNSF-HA увеличивал его (рис. 2h, i). Этот результат предполагает, что очень большие молекулы vHMM-HA экранируют CD44 и уменьшают его взаимодействие с другими белками.
2g). Эти результаты показывают, что белок-белковые взаимодействия CD44 стимулируются HMM-HA, но подавляются vHMM-HA. Затем, чтобы прояснить влияние vHMM-HA на белковое взаимодействие CD44-CD44, мы одновременно сверхэкспрессировали CD44-FLAG и CD44-YFP в IMR9.0 клеток и исследовали их взаимодействие с помощью анализа коиммунопреципитации FLAG. В соответствии с нашими данными масс-спектрометрии, cNSF-HA уменьшал количество коиммунопреципитированного CD44-YFP, тогда как fNSF-HA увеличивал его (рис. 2h, i). Этот результат предполагает, что очень большие молекулы vHMM-HA экранируют CD44 и уменьшают его взаимодействие с другими белками.
vHMM- и HMM-HA оказывают противоположное влияние на экспрессию генов
Наши данные показывают, что vHMM-HA защищает клетки, повышая устойчивость клеток к стрессу посредством измененной передачи сигналов CD44. В подтверждение этого мнения, предварительная инкубация с NSF-HA не снижала индукцию внутриклеточных АФК после обработки tBHP у IMR9.0 ячеек (дополнительный рис. 5A). Кроме того, непродолжительное лечение HAase после инкубации NSF-HA не ухудшило его цитозащитный эффект (дополнительная рис. 5B, C). Это указывает на то, что остаточная перицеллюлярная ГК вряд ли важна для цитопротекции. Хотя интернализованный HA может быть вовлечен, наши данные, тем не менее, предполагают, что внутриклеточные сигналы ниже CD44 играют ключевую роль в цитопротекции с помощью vHMM-HA.
5A). Кроме того, непродолжительное лечение HAase после инкубации NSF-HA не ухудшило его цитозащитный эффект (дополнительная рис. 5B, C). Это указывает на то, что остаточная перицеллюлярная ГК вряд ли важна для цитопротекции. Хотя интернализованный HA может быть вовлечен, наши данные, тем не менее, предполагают, что внутриклеточные сигналы ниже CD44 играют ключевую роль в цитопротекции с помощью vHMM-HA.
Таким образом, для дальнейшего изучения механизмов цитопротекторного эффекта мы сначала стремились выяснить, как vHMM-HA влияет на сигнатуру транскрипции в стрессовых клетках. Мы сравнили транскриптомы IMR90 клеток через 6 ч после начала 1-часовой обработки низкой дозой tBHP и идентифицировали 745 генов, которые по-разному экспрессируются между предварительно инкубированными клетками cNSF-HA – и fNSF-HA ( q < 0,05; далее обозначаемые как HA гены, зависящие от длины полимера). Уровни экспрессии большинства этих генов противоположно регулировались cNSF-HA и fNSF-HA (достоверная отрицательная корреляция; тест корреляции Спирмена, p < 2,2 × 10 -16 ) (рис. 3a), как и в случае с Белок-белковые взаимодействия CD44. Таким образом, хотя ИПП CD44 исследовали в условиях сверхэкспрессии CD44, которая может повлиять на результаты, наши протеомный и транскриптомный анализы показали противоположные эффекты cNSF-HA и fNSF-HA. Анализ генетической онтологии 26 генов, зависящих от длины полимера HA, показали обогащение генами, кодирующими регуляторы и интеракторы p53 (рис. 3b). Затем мы провели анализ обогащения сайта связывания фактора транскрипции (TFBS) на основе мотива 27 и обнаружили, что мишени регулируемых CD44 факторов транскрипции, таких как ELK1 28 и EGR1 29 , были чрезмерно представлены в генах, зависящих от длины полимера HA. (рис. 3в). Более того, было показано, что наиболее обогащенные мишени фактора транскрипции, а именно мишени HIF1α, значительно перекрываются с мишенями CD44-ICD (внутрицитоплазматический домен CD44), который образуется в результате протеолиза CD44 30 . Действительно, уровни CD44-ICD были ниже в клетках, инкубированных с fNSF-HA (дополнительная фигура 6), и наблюдалось обогащение мотива связывания CD44-ICD (CCTGCG) в промоторных областях размером 500 п.
3a), как и в случае с Белок-белковые взаимодействия CD44. Таким образом, хотя ИПП CD44 исследовали в условиях сверхэкспрессии CD44, которая может повлиять на результаты, наши протеомный и транскриптомный анализы показали противоположные эффекты cNSF-HA и fNSF-HA. Анализ генетической онтологии 26 генов, зависящих от длины полимера HA, показали обогащение генами, кодирующими регуляторы и интеракторы p53 (рис. 3b). Затем мы провели анализ обогащения сайта связывания фактора транскрипции (TFBS) на основе мотива 27 и обнаружили, что мишени регулируемых CD44 факторов транскрипции, таких как ELK1 28 и EGR1 29 , были чрезмерно представлены в генах, зависящих от длины полимера HA. (рис. 3в). Более того, было показано, что наиболее обогащенные мишени фактора транскрипции, а именно мишени HIF1α, значительно перекрываются с мишенями CD44-ICD (внутрицитоплазматический домен CD44), который образуется в результате протеолиза CD44 30 . Действительно, уровни CD44-ICD были ниже в клетках, инкубированных с fNSF-HA (дополнительная фигура 6), и наблюдалось обогащение мотива связывания CD44-ICD (CCTGCG) в промоторных областях размером 500 п. н. генов, зависящих от длины полимера HA. (рис. 3d).
н. генов, зависящих от длины полимера HA. (рис. 3d).
a Гены, дифференциально экспрессирующиеся в клетках IMR90 при обработке cNSF-HA или fNSF-HA (гены, зависящие от длины полимера HA). Клетки предварительно инкубировали в течение 6 часов с 20 мкг/мл cNSF-HA, fNSF-HA или PBS, затем удаляли HA и клетки подвергали воздействию 200 мкМ tBHP в течение 1 часа. Клетки собирали через 6 ч после начала обработки tBHP. b Обогащение GO Term, пути Reactome и белок-белковых взаимодействий фактора транскрипции (PPI) в генах, зависящих от длины полимера HA. c , d Обогащение сайтов связывания факторов транскрипции (TFBS) в генах, зависящих от длины полимера HA. Белые столбцы представляют собой обогащение TFBS всеми генами, зависящими от длины полимера HA. Серые столбцы представляют собой обогащение TFBS генами, зависящими от длины полимера HA, которые участвуют в регуляции транскрипции с помощью p53 или кодируют белки, взаимодействующие с p53. Черные столбцы представляют собой обогащение TFBS генами-мишенями p53, зависящими от длины полимера HA. e Дифференциальные уровни экспрессии генов, зависящих от длины полимера HA (определяемых в присутствии tBHP), в клетках IMR90, инкубированных в течение 6 ч с 20 мкг/мл cNSF-HA, fNSF-HA или PBS. f Скрипичный график представляет собой распределение p -значений двустороннего t -критерия, сравнивающего уровни экспрессии генов, зависящих от длины полимера HA, и генов, независимых от длины полимера HA (определено в наличие tBHP) между cNSF-HA- и fNSF-HA-инкубированным IMR90 ячеек. Клетки инкубировали в течение 6 ч с 20 мкг/мл cNSF-HA или fNSF-HA без дополнительной обработки tBHP. г Дифференциальные уровни экспрессии генов, зависящих от длины полимера HA (определялись в присутствии tBHP) в контрольных и сверхэкспрессирующих CD44 клетках IMR90, инкубированных в течение 6 ч с 20 мкг/мл cNSF-HA или PBS.
Черные столбцы представляют собой обогащение TFBS генами-мишенями p53, зависящими от длины полимера HA. e Дифференциальные уровни экспрессии генов, зависящих от длины полимера HA (определяемых в присутствии tBHP), в клетках IMR90, инкубированных в течение 6 ч с 20 мкг/мл cNSF-HA, fNSF-HA или PBS. f Скрипичный график представляет собой распределение p -значений двустороннего t -критерия, сравнивающего уровни экспрессии генов, зависящих от длины полимера HA, и генов, независимых от длины полимера HA (определено в наличие tBHP) между cNSF-HA- и fNSF-HA-инкубированным IMR90 ячеек. Клетки инкубировали в течение 6 ч с 20 мкг/мл cNSF-HA или fNSF-HA без дополнительной обработки tBHP. г Дифференциальные уровни экспрессии генов, зависящих от длины полимера HA (определялись в присутствии tBHP) в контрольных и сверхэкспрессирующих CD44 клетках IMR90, инкубированных в течение 6 ч с 20 мкг/мл cNSF-HA или PBS.
Изображение в полный размер
Все эти обогащения TFBS становятся более заметными, когда анализ был ограничен генами, зависящими от длины полимера HA, кодирующими регуляторы и интеракторы p53 (рис. 3d), что подразумевает основную роль пути p53 в CD44-зависимый цитопротекторный эффект NSF-HA. В соответствии с этими наблюдениями произошло значительное обогащение курируемых высоконадежных генов-мишеней p53 31 в генах, зависящих от длины полимера HA (точный критерий Фишера, p = 1,7 × 10 -6 ) (дополнительный рисунок 7A). Примечательно, что предварительная инкубация cNSF-HA подавляла экспрессию большинства генов-мишеней p53, зависящих от длины полимера HA (дополнительная фиг. 7A, B). При этом большинство генов-мишеней p53 одинаково реагируют на окислительный стресс в клетках cNSF-HA, fNSF-HA и PBS (дополнительная рис. 7C). HA также не влияла на цитоплазматический и ядерный уровни p53 (дополнительная рис. 7D – G). Гены-мишени CD44-ICD и CD44-регулируемых факторов транскрипции не были обогащены генами-мишенями p53, зависящими от длины полимера HA (рис. 3d), что позволяет предположить, что транскрипционная модуляция генов-мишеней p53, зависящих от длины полимера HA, является следствием измененных экспрессия генов, кодирующих регуляторы и интеракторы р53, которые регулируются CD44-регулируемыми транскрипционными факторами и CD44-ICD.
3d), что подразумевает основную роль пути p53 в CD44-зависимый цитопротекторный эффект NSF-HA. В соответствии с этими наблюдениями произошло значительное обогащение курируемых высоконадежных генов-мишеней p53 31 в генах, зависящих от длины полимера HA (точный критерий Фишера, p = 1,7 × 10 -6 ) (дополнительный рисунок 7A). Примечательно, что предварительная инкубация cNSF-HA подавляла экспрессию большинства генов-мишеней p53, зависящих от длины полимера HA (дополнительная фиг. 7A, B). При этом большинство генов-мишеней p53 одинаково реагируют на окислительный стресс в клетках cNSF-HA, fNSF-HA и PBS (дополнительная рис. 7C). HA также не влияла на цитоплазматический и ядерный уровни p53 (дополнительная рис. 7D – G). Гены-мишени CD44-ICD и CD44-регулируемых факторов транскрипции не были обогащены генами-мишенями p53, зависящими от длины полимера HA (рис. 3d), что позволяет предположить, что транскрипционная модуляция генов-мишеней p53, зависящих от длины полимера HA, является следствием измененных экспрессия генов, кодирующих регуляторы и интеракторы р53, которые регулируются CD44-регулируемыми транскрипционными факторами и CD44-ICD.
Интересно, что мы обнаружили, что ранние изменения экспрессии генов, чувствительных к стрессу, индуцированные tBHP и облучением 32 , показывают положительную корреляцию с изменениями экспрессии генов, индуцированными предварительной инкубацией cNSF-HA в генах, зависящих от длины полимера HA (критерий корреляции Спирмена, ). p < 2,2 × 10 −16 для tBHP и p = 3,2 × 10 −5 для облучения, дополнительный рисунок 7H). Это удивительно, поскольку гены-мишени p53 в целом преимущественно активировались при этих ранних реакциях на стресс (дополнительная фигура 7A). Эти результаты свидетельствуют о том, что предварительная инкубация cNSF-HA потенцирует подмножество ранней реакции на стресс, которая особенно связана с супрессией p53.
Важно отметить, что транскриптомный анализ не подвергнутых стрессу клеток IMR90, инкубированных с cNSF-HA, fNSF-HA и PBS, показал, что изменения экспрессии большинства генов, зависящих от длины полимера HA, уже индуцировались до обработки tBHP (рис. 3e). ). Другие гены показали гораздо меньшие эффекты, зависящие от длины полимера HA, в этих клетках (рис. 3f). RT-qPCR анализ клеток IMR90 без стресса и CD44-нокдауна подтвердил, что зависящие от длины полимера эффекты HA на экспрессию генов-мишеней p53 ( RRM2B 33 , PRRX2 25 и PYCARD 34 ) опосредуются CD44 (дополнительный рисунок 8). Наконец, чтобы дополнительно подтвердить роль передачи сигналов CD44 в регуляции генов, зависящих от длины полимера HA (и в дополнение к протеомному анализу, описанному в предыдущем разделе), мы провели транскриптомный анализ ложно- и CD44-сверхэкспрессирующих клеток IMR90 под нестрессовые состояния. Как и ожидалось на основании вышеприведенного протеомного анализа, транскрипционный эффект сверхэкспрессии CD44 на гены, зависящие от длины полимера HA, в значительной степени подавлялся при инкубации клеток с cNSF-HA. В генах, зависящих от длины полимера HA, изменения экспрессии, вызванные сверхэкспрессией CD44, в IMR9, обработанном PBS0 и клетки, индуцированные обработкой cNSF-HA в клетках IMR90 со сверхэкспрессией CD44, показали значительную отрицательную корреляцию (корреляционный тест Спирмена, p < 2,2 × 10 -16 ) (рис.
3e). ). Другие гены показали гораздо меньшие эффекты, зависящие от длины полимера HA, в этих клетках (рис. 3f). RT-qPCR анализ клеток IMR90 без стресса и CD44-нокдауна подтвердил, что зависящие от длины полимера эффекты HA на экспрессию генов-мишеней p53 ( RRM2B 33 , PRRX2 25 и PYCARD 34 ) опосредуются CD44 (дополнительный рисунок 8). Наконец, чтобы дополнительно подтвердить роль передачи сигналов CD44 в регуляции генов, зависящих от длины полимера HA (и в дополнение к протеомному анализу, описанному в предыдущем разделе), мы провели транскриптомный анализ ложно- и CD44-сверхэкспрессирующих клеток IMR90 под нестрессовые состояния. Как и ожидалось на основании вышеприведенного протеомного анализа, транскрипционный эффект сверхэкспрессии CD44 на гены, зависящие от длины полимера HA, в значительной степени подавлялся при инкубации клеток с cNSF-HA. В генах, зависящих от длины полимера HA, изменения экспрессии, вызванные сверхэкспрессией CD44, в IMR9, обработанном PBS0 и клетки, индуцированные обработкой cNSF-HA в клетках IMR90 со сверхэкспрессией CD44, показали значительную отрицательную корреляцию (корреляционный тест Спирмена, p < 2,2 × 10 -16 ) (рис. 3g). В совокупности эти данные показывают, что vHMM-HA противодействует транскрипционному эффекту HMM-HA и CD44 и, таким образом, изменяет экспрессию интеракторов и регуляторов p53, которые потенциально находятся под контролем факторов транскрипции, связанных с CD44, что приводит к снижению экспрессии подмножества генов-мишеней p53.
3g). В совокупности эти данные показывают, что vHMM-HA противодействует транскрипционному эффекту HMM-HA и CD44 и, таким образом, изменяет экспрессию интеракторов и регуляторов p53, которые потенциально находятся под контролем факторов транскрипции, связанных с CD44, что приводит к снижению экспрессии подмножества генов-мишеней p53.
vHMM-HA защищает клетки посредством ослабления p53
Затем мы попытались выяснить, влияют ли vHMM-HA и HMM-HA по-разному на статус самого p53. Мы исследовали основные сайты фосфорилирования N-конца p53 и обнаружили, что p53 Ser9 был менее фосфорилирован в клетках, инкубированных с cNSF-HA, чем в клетках, инкубированных с fNSF-HA (рис. 4a, b и дополнительная рис. 9). Чтобы проверить, действительно ли p53 участвует в цитопротекторном эффекте NSF-HA, мы сбили p53 в клетках IMR90, используя проверенную миРНК 9.0009 35 . Клетки IMR90, в которых был нокдаун p53, все еще были чувствительны к tBHP, но больше не были защищены предварительной инкубацией NSF-HA (рис. 4c–e). Поскольку siRNA не может полностью заглушить ген-мишень, мы затем провели тот же эксперимент с использованием MSF 36 с нокаутом p53 дикого типа. Подобно клеткам IMR90, MSF с нокаутом p53 все еще были чувствительны к tBHP, но не были защищены предварительной инкубацией NSF-HA (рис. 4f–h). Основываясь на этих результатах, мы делаем вывод, что цитопротекторный эффект NSF-HA опосредуется, по крайней мере, частично за счет ослабления пути p53.
4c–e). Поскольку siRNA не может полностью заглушить ген-мишень, мы затем провели тот же эксперимент с использованием MSF 36 с нокаутом p53 дикого типа. Подобно клеткам IMR90, MSF с нокаутом p53 все еще были чувствительны к tBHP, но не были защищены предварительной инкубацией NSF-HA (рис. 4f–h). Основываясь на этих результатах, мы делаем вывод, что цитопротекторный эффект NSF-HA опосредуется, по крайней мере, частично за счет ослабления пути p53.
a Вестерн-блоты, показывающие уровни p-p53-Ser9 в клетках IMR90, обработанных 20 мкг/мл cNSF-HA или fNSF-HA в течение 6 часов. b Количественная оценка Вестерн-блотов показана в ( a ) на изображении J. Средние значения взяты из трех биологических повторов. Для каждой биологической повторности иммуноблотинг проводили трижды и усредняли. c Нокдаун p53 отменяет цитопротекторный эффект NSF-HA на гибель клеток, вызванную окислительным стрессом, в IMR90 ячеек. Процент живых клеток измеряли путем окрашивания аннексином-V/PI через 1 день после 1 часа обработки 3 мМ tBHP ( n = 3). Клетки предварительно инкубировали в течение 6 ч с 20 мкг/мл NSF-HA или PBS перед обработкой tBHP. siRNA трансфицировали за 2 дня до первой обработки tBHP. d p53-нокдаун отменяет цитопротекторный эффект NSF-HA на индуцированную окислительным стрессом остановку роста в клетках IMR90 ( n = 4). Клетки предварительно инкубировали в течение 6 ч с 20 мкг/мл NSF-HA или PBS в течение 6 ч, затем ГК удаляли и клетки подвергали воздействию 200 мкМ tBHP в течение 1 ч в день 0 и день 1. миРНК трансфицировали за 2 дня до день 0. e Подтверждение нокдауна p53 в клетках IMR90 вестерн-блоттингом. Клетки собирали через 2 дня после трансфекции р53 или контрольной миРНК. Иммуноблотинг был повторен один раз с аналогичным результатом. f Нокаут p53 отменяет цитопротекторный эффект NSF-HA на гибель клеток MSF, вызванную окислительным стрессом. Процентное содержание живых MSF измеряли путем окрашивания аннексином-V/PI через 1 день после 1 часа обработки 1,5 мМ tBHP ( n = 3).
Процент живых клеток измеряли путем окрашивания аннексином-V/PI через 1 день после 1 часа обработки 3 мМ tBHP ( n = 3). Клетки предварительно инкубировали в течение 6 ч с 20 мкг/мл NSF-HA или PBS перед обработкой tBHP. siRNA трансфицировали за 2 дня до первой обработки tBHP. d p53-нокдаун отменяет цитопротекторный эффект NSF-HA на индуцированную окислительным стрессом остановку роста в клетках IMR90 ( n = 4). Клетки предварительно инкубировали в течение 6 ч с 20 мкг/мл NSF-HA или PBS в течение 6 ч, затем ГК удаляли и клетки подвергали воздействию 200 мкМ tBHP в течение 1 ч в день 0 и день 1. миРНК трансфицировали за 2 дня до день 0. e Подтверждение нокдауна p53 в клетках IMR90 вестерн-блоттингом. Клетки собирали через 2 дня после трансфекции р53 или контрольной миРНК. Иммуноблотинг был повторен один раз с аналогичным результатом. f Нокаут p53 отменяет цитопротекторный эффект NSF-HA на гибель клеток MSF, вызванную окислительным стрессом. Процентное содержание живых MSF измеряли путем окрашивания аннексином-V/PI через 1 день после 1 часа обработки 1,5 мМ tBHP ( n = 3). Клетки предварительно инкубировали в течение 6 ч с 20 мкг/мл NSF-HA или PBS перед обработкой tBHP. г p53-нокаут отменяет цитопротекторный эффект NSF-HA на индуцированную окислительным стрессом остановку роста MSF ( n = 4). Клетки предварительно инкубировали в течение 6 ч с 20 мкг/мл NSF-HA или PBS в течение 6 ч, затем ГК удаляли и клетки подвергали воздействию 150 мкМ tBHP в течение 1 ч в день 0 и день 1. ч Подтверждение нокаута p53 в MSF с помощью вестерн-блоттинга. Иммуноблотинг был повторен один раз с аналогичным результатом. Столбики погрешностей представлены в виде средних значений ± SD. * p < 0,05 (двусторонний т -тест).
Клетки предварительно инкубировали в течение 6 ч с 20 мкг/мл NSF-HA или PBS перед обработкой tBHP. г p53-нокаут отменяет цитопротекторный эффект NSF-HA на индуцированную окислительным стрессом остановку роста MSF ( n = 4). Клетки предварительно инкубировали в течение 6 ч с 20 мкг/мл NSF-HA или PBS в течение 6 ч, затем ГК удаляли и клетки подвергали воздействию 150 мкМ tBHP в течение 1 ч в день 0 и день 1. ч Подтверждение нокаута p53 в MSF с помощью вестерн-блоттинга. Иммуноблотинг был повторен один раз с аналогичным результатом. Столбики погрешностей представлены в виде средних значений ± SD. * p < 0,05 (двусторонний т -тест).
Изображение полного размера
Обсуждение
Хотя внеклеточный матрикс составляет основную часть тела, его связь со старением остается в значительной степени неизученной. HA, линейный полисахарид и основной компонент внеклеточного матрикса, имеет очень высокую молекулярную массу у самого долгоживущего грызуна, NMR, но было неясно, способствует ли исключительная длина полимера vHMM-HA фенотипам NMR. Здесь мы продемонстрировали, что vHMM-HA (> 6,1 MDa) функционально отличается от более короткой высокомолекулярной HA и что vHMM-HA проявляет отчетливое цитопротекторное действие на клетки ЯМР, мыши и человека в зависимости от р53. образом. Наши данные в совокупности подтверждают модель, в которой vHMM-HA подавляет белок-белковые взаимодействия и передачу сигналов с помощью CD44, что приводит к изменению экспрессии интеракторов, регуляторов и мишеней р53 и, таким образом, проявляет отчетливую цитопротекцию посредством модулирования пути р53 (рис. 5). Наши результаты показывают, что vHMM-HA способствует долговечности ЯМР, и подчеркивают потенциальное применение vHMM-HA в качестве цитозащитной молекулы.
Здесь мы продемонстрировали, что vHMM-HA (> 6,1 MDa) функционально отличается от более короткой высокомолекулярной HA и что vHMM-HA проявляет отчетливое цитопротекторное действие на клетки ЯМР, мыши и человека в зависимости от р53. образом. Наши данные в совокупности подтверждают модель, в которой vHMM-HA подавляет белок-белковые взаимодействия и передачу сигналов с помощью CD44, что приводит к изменению экспрессии интеракторов, регуляторов и мишеней р53 и, таким образом, проявляет отчетливую цитопротекцию посредством модулирования пути р53 (рис. 5). Наши результаты показывают, что vHMM-HA способствует долговечности ЯМР, и подчеркивают потенциальное применение vHMM-HA в качестве цитозащитной молекулы.
Наши данные показывают, что vHMM-HA ослабляет белок-белковые взаимодействия CD44 (PPI), тогда как HMM-HA их усиливает. Одновременно vHMM-HA подавляет экспрессию CD44-зависимых генов, в том числе регулируемых CD44-зависимыми факторами транскрипции, такими как ELK, EGR1 и CD44-ICD. Таким образом, vHMM-HA и HMM-HA вызывают противоположные эффекты на экспрессию CD44-зависимых генов, которые также связаны с путем р53. В результате vHMM-HA частично ослабляет р53 и его гены-мишени и защищает клетки р53-зависимым образом.
Таким образом, vHMM-HA и HMM-HA вызывают противоположные эффекты на экспрессию CD44-зависимых генов, которые также связаны с путем р53. В результате vHMM-HA частично ослабляет р53 и его гены-мишени и защищает клетки р53-зависимым образом.
Изображение в натуральную величину
Следует отметить, однако, что наши механические исследования были в значительной степени ограничены клетками человека как клинически значимой моделью. Гены, участвующие в механизмах, опосредующих защитный эффект vHMM-HA, могут различаться у разных видов. Например, существуют значительные различия в генах, регулируемых p53, между человеком и мышью 37 . Неудивительно, что аминокислотные последовательности р53 различаются у человека, мыши и ЯМР 38 , хотя о функциональных различиях между NMR и двумя другими видами не сообщалось. Еще одним важным ограничением нашего исследования является то, что мы сосредоточились только на отличительном цитопротекторном эффекте vHMM-HA. Другие функции также могут различаться между vHMM-HA и более коротким HMM-HA. Кроме того, в то время как сигналы CD44-p53 играют критическую роль в цитозащитном эффекте vHMM-HA, другие HA-связывающие белки, такие как versican 39 , а также другие аспекты CD44, такие как образование молекулярных комплексов 40 и частокол 41 , могут быть вовлечены в цитопротекторный эффект или другую функцию vHMM-HA. Хотя эти вопросы выходят за рамки данного исследования, наши протеомные и транскриптомные данные могут быть полезными ресурсами для решения этих вопросов.
Кроме того, в то время как сигналы CD44-p53 играют критическую роль в цитозащитном эффекте vHMM-HA, другие HA-связывающие белки, такие как versican 39 , а также другие аспекты CD44, такие как образование молекулярных комплексов 40 и частокол 41 , могут быть вовлечены в цитопротекторный эффект или другую функцию vHMM-HA. Хотя эти вопросы выходят за рамки данного исследования, наши протеомные и транскриптомные данные могут быть полезными ресурсами для решения этих вопросов.
В текущем исследовании мы в основном использовали ГК естественного происхождения, которые имеют значительную полидисперсность, и, таким образом, может быть функциональная гетерогенность даже внутри vHMM-HA и HMM-HA. Однако, основываясь на результатах наших экспериментов с использованием гель-экстрагированного vHMM-HA и 1 МДа Select-HA TM , мы можем заключить, что vHMM-HA с молекулярной массой более 6,1 МДа обладает превосходным защитным эффектом по сравнению с 1 МДа HA или MSF-HA, состоящим из HMM-HA с молекулярной массой менее 6,1 МДа. Вывод о том, что NSF-HA обладает лучшим цитопротекторным эффектом по сравнению с MSF-HA, также верен. Эти выводы предполагают вклад vHMM-HA в стрессоустойчивость и долговечность ЯМР.
Вывод о том, что NSF-HA обладает лучшим цитопротекторным эффектом по сравнению с MSF-HA, также верен. Эти выводы предполагают вклад vHMM-HA в стрессоустойчивость и долговечность ЯМР.
Хотя vHMM-HA частично ослабляет активность опухолевого супрессора p53, ЯМР, тем не менее, чрезвычайно устойчив к канцерогенезу. Одно из объяснений состоит в том, что vHMM-HA-индуцированная частичная аттенуация р53 не обязательно может ставить под угрозу его супрессорную активность. Действительно, в целом гены-мишени p53 очень сходно реагируют на окислительный стресс в клетках, инкубированных с PBS и HA. Дифференциальная модуляция транскрипции подмножества генов-мишеней p53 с помощью cNSF-HA и fNSF-HA может быть следствием измененной экспрессии генов, кодирующих регуляторы и интеракторы p53, которые регулируются CD44-регулируемыми транскрипционными факторами и CD44-ICD.
Наше наблюдение, что на физиологических уровнях vHMM-HA лучше защищает клетки человека от стресса, чем более короткие HMM-HA, имеет важное клиническое значение, учитывая широкое медицинское применение HA 42 . Производство ГК с более высокой молекулярной массой может привести к улучшению медицинских продуктов на основе ГК.
Производство ГК с более высокой молекулярной массой может привести к улучшению медицинских продуктов на основе ГК.
Методы
Культура клеток
NSF и MSF были выделены в нашей лаборатории 43 из кожи молодых взрослых мышей NMR и C57BL/6J, соответственно. Фибробласты кожи выделяли из образцов кожи, взятых у умерщвленных животных, путем измельчения и переваривания их с помощью 0,14 единиц Вунча/мл Liberase TM (Roche Diagnostics) в среде DMEM/F12 при 37°С в течение 60 мин. Все животные содержались в соответствии с правилами, установленными и одобренными Комитетом по животным ресурсам Университета Рочестера (UCAR), который придерживается рекомендаций FDA и NIH по уходу за животными и проверяет все протоколы животных до утверждения. IMR90 фибробластов легких человека были получены из Института медицинских исследований Кориелла. Клетки HEK293T были получены от ATCC. Клетки NSF, MSF и IMR90 культивировали в среде EMEM с добавлением 15% FBS. NSF культивировали при 32 °C, 4,8% CO 2 , 2,8% O 2 . Клетки MSF и IMR90 культивировали при 37 °C, 5% CO 2 , 4% O 2 . Количество пассажей клеток NSF, MSF и IMR90 составляло менее 20, 5 и 40 соответственно. В клетках IMR90 последовательность всех экзонов p53 была проверена с использованием наших данных RNA-Seq, и было обнаружено отсутствие мутаций. Подсчет клеток проводили с использованием культуральных чашек с сеткой.
Клетки MSF и IMR90 культивировали при 37 °C, 5% CO 2 , 4% O 2 . Количество пассажей клеток NSF, MSF и IMR90 составляло менее 20, 5 и 40 соответственно. В клетках IMR90 последовательность всех экзонов p53 была проверена с использованием наших данных RNA-Seq, и было обнаружено отсутствие мутаций. Подсчет клеток проводили с использованием культуральных чашек с сеткой.
Анализ апоптоза
Количественное определение апоптотических клеток проводили с использованием набора для обнаружения апоптоза аннексин-V (eBioscience) в соответствии с инструкциями производителя. После окрашивания клетки анализировали на проточном цитометре LSR-II (BD Biosciences).
Препарат ГК
Для очистки ГК кондиционированные среды сначала смешивали с раствором протеиназы К (конечные концентрации 1 мМ Трис-Cl, рН 8,0, 2,5 мМ ЭДТА, 10 мМ NaCl, 0,05% SDS, 1 мг/мл протеиназы К) и инкубировали при 55°С в течение 4 ч. После расщепления белка среду экстрагировали насыщенным раствором фенол-хлороформ-изоамиловый спирт (Sigma). ГК осаждали этанолом и центрифугированием (4000× г в течение 45 мин). Осадок ГК растворяли в PBS и затем экстрагировали 1/100 объема Triton-X114 44 . После экстракции Тритоном-Х114 ГК снова осаждали этанолом. Наконец, осадок ГК промывали 70% этанолом и растворяли в PBS. Для гель-экстракции vHMM-HA NSF-HA использовали на 0,7% легкоплавкой агарозе (бляшка Agarose-LM; Nacalai Tesque) в буфере TBE. Маркерную дорожку отрезали от геля и окрашивали, как описано ниже. Гель, содержащий NSF-HA, с маркерной полосой выше 6,1 MДа, вырезали, и vHMM-HA выделяли из геля с помощью термостабильной агаразы (ген Nippon) в соответствии с инструкциями производителя. Восстановленный vHMM-HA затем экстрагировали PCI и Triton-X114 и осаждали этанолом, как описано выше. Концентрацию ГК измеряли с помощью карбазольного анализа 45 . Streptomyces HAase (Sigma) использовали для деградации HA в кондиционированных средах и для частичной фрагментации HA. HA частично фрагментировали путем инкубации с 0,5 ед/мл HAase при 37°C в течение 60 мин с последующей тепловой инактивацией при 95°C в течение 10 мин.
ГК осаждали этанолом и центрифугированием (4000× г в течение 45 мин). Осадок ГК растворяли в PBS и затем экстрагировали 1/100 объема Triton-X114 44 . После экстракции Тритоном-Х114 ГК снова осаждали этанолом. Наконец, осадок ГК промывали 70% этанолом и растворяли в PBS. Для гель-экстракции vHMM-HA NSF-HA использовали на 0,7% легкоплавкой агарозе (бляшка Agarose-LM; Nacalai Tesque) в буфере TBE. Маркерную дорожку отрезали от геля и окрашивали, как описано ниже. Гель, содержащий NSF-HA, с маркерной полосой выше 6,1 MДа, вырезали, и vHMM-HA выделяли из геля с помощью термостабильной агаразы (ген Nippon) в соответствии с инструкциями производителя. Восстановленный vHMM-HA затем экстрагировали PCI и Triton-X114 и осаждали этанолом, как описано выше. Концентрацию ГК измеряли с помощью карбазольного анализа 45 . Streptomyces HAase (Sigma) использовали для деградации HA в кондиционированных средах и для частичной фрагментации HA. HA частично фрагментировали путем инкубации с 0,5 ед/мл HAase при 37°C в течение 60 мин с последующей тепловой инактивацией при 95°C в течение 10 мин. Select-HA TM был от Echelon Biosciences. Для блокирования взаимодействий HA/CD44 в культуральную среду добавляли нейтрализующее антитело к CD44 (Clone 2C5; R&D) в концентрации 10 нг/мл.
Select-HA TM был от Echelon Biosciences. Для блокирования взаимодействий HA/CD44 в культуральную среду добавляли нейтрализующее антитело к CD44 (Clone 2C5; R&D) в концентрации 10 нг/мл.
Экстракция РНК, обратная транскрипция и количественная ПЦР в реальном времени
Тотальную РНК экстрагировали с помощью RNAiso Plus (Takara) и подвергали обратной транскрипции в кДНК со случайными гексамерами с использованием набора реагентов PrimeScript (Takara). КПЦР в реальном времени проводили с использованием TB Green TM Premix Ex Taq II TM (Takara) с системой ПЦР в реальном времени Step ONE Plus (Life Technologies). В этом исследовании использовались следующие праймеры: ACTB вперед, AGATCAAGATCATTGCTCCTCCTG; АКТВ реверс GCCGGACTCGTCATACTCCT; CD44 вперед, TGGTGAACAAGGAGTCGTCA; CD44 реверс, ACACCCCAATCTTCATGTCC; PRRX2 вперед, AGGTGCCTACGGTGAACTGA; PRRX2 реверс, CTGCCCCCTTTTCTATTGCT; PYCARD вперед, TGACGGATGAGCAGTACCAG; PYCARD реверс, CAGGCTGGTGTGTGAAACTGAA; RHAMM вперед, AGCAAGAAGGCATGGAGATG; RHAMM реверс, CCCTCCAGTTGGGCTATTTT; RRM2B вперед, GCCAGGACTCACTTTTTTCCA; РРМ2Б реверс, TCCCTGACCCTTTCTTCTGA.
Гель-электрофорез в импульсном поле
Очищенную ГК смешивали с раствором сахарозы (конечная концентрация 333 мМ) и загружали в 0,4% агарозный гель SeaKem Gold (Lonza). Вместе с образцами запускали HA-Ladders (Hyalose). Образцы анализировали в течение 12 ч при 4°С и 75°В с рабочим соотношением 1–10 в буфере ТВЕ с использованием системы CHEF-DRII (Bio-Rad). После анализа гель окрашивали 0,005% (масса/объем) красителем Stains-All (Santa Cruz) в 50% этаноле в течение ночи. Затем гель дважды промывали 10% этанолом, подвергали воздействию света для уменьшения фона и фотографировали с помощью ChemiDoc Imaging System (Bio-Rad).
Иммунофлуоресцентное окрашивание
Клетки промывали PBS и фиксировали 4% формальдегидом при комнатной температуре в течение 15 мин. После фиксации клетки промывали 3% BSA/PBS и пермеабилизировали 0,5% Triton X-100/PBS при комнатной температуре в течение 20 мин. Затем клетки промывали 3% BSA/PBS и блокировали блокирующим буфером (10% FBS и 1% BSA в PBS) при комнатной температуре в течение 1 часа. Затем клетки инкубировали с антителами γh3AX (JBW301; Millipore; 1:500) и 53BP1 (ab172580; Abcam; 1:500) в блокирующем буфере при 4 °C в течение ночи и промывали PBST. Наконец, клетки инкубировали с козьим антимышиным IgG Alexa Fluor 488 и козьим антикроличьи IgG Alexa Fluor 568 (Thermo Fisher Scientific; 1:1000) в блокирующем буфере при комнатной температуре в течение 1 часа, промывали PBST и помещали в Vectashield. содержащий DAPI (Vector Laboratories). Изображения были получены с использованием конфокального микроскопа Leica SP5.
Затем клетки инкубировали с антителами γh3AX (JBW301; Millipore; 1:500) и 53BP1 (ab172580; Abcam; 1:500) в блокирующем буфере при 4 °C в течение ночи и промывали PBST. Наконец, клетки инкубировали с козьим антимышиным IgG Alexa Fluor 488 и козьим антикроличьи IgG Alexa Fluor 568 (Thermo Fisher Scientific; 1:1000) в блокирующем буфере при комнатной температуре в течение 1 часа, промывали PBST и помещали в Vectashield. содержащий DAPI (Vector Laboratories). Изображения были получены с использованием конфокального микроскопа Leica SP5.
Получение клеток IMR90 со сверхэкспрессией CD44
Клетки IMR90 со сверхэкспрессией CD44 и CD44-FLAG получали с помощью вирусной трансдукции и отбора 1 мкг/мл пуромицина. Клетки IMR90 со сверхэкспрессией CD44-FLAG дополнительно трансдуцировали ретровирусным вектором, несущим CD44-YFP, и отбирали с помощью FACS (Sony SH800). Ретровирусный супернатант готовили путем совместной трансфекции клеток HEK293T векторами VSV-G, pUMVC и pBabe-puro. Пустые и экспрессирующие CD44s векторы pBabe были получены от Addgene (#1764 и #19). 127). CD44 были клонированы в pcDNA3.1-ZNF598-TEV-3xFLAG (#105690, Addgene) и pcDNA3-YFP (#13033, Addgene) для получения CD44-FLAG (C-конец) и CD44-YFP (C-конец) соответственно. . Затем CD44-FLAG и CD44-YFP клонировали обратно в вектор pBabe для получения вируса.
127). CD44 были клонированы в pcDNA3.1-ZNF598-TEV-3xFLAG (#105690, Addgene) и pcDNA3-YFP (#13033, Addgene) для получения CD44-FLAG (C-конец) и CD44-YFP (C-конец) соответственно. . Затем CD44-FLAG и CD44-YFP клонировали обратно в вектор pBabe для получения вируса.
Иммунопреципитация
Клетки инкубировали с 20 мкг/мл cNSF-HA, fNSF-HA или эквивалентным объемом PBS в течение 6 ч, затем среду удаляли и клетки сшивали в PBS, содержащем 500 мкМ DSP и 500 мкМ DTME (Thermo Fisher Scientific) в течение 30 мин при комнатной температуре. Сшивку гасили инкубацией с PBS, содержащим 20 мМ Tris-Cl (pH 7,4) и 5 мМ L-цистеина (Sigma). Клетки собирали в буфере RIPA (50 мМ Трис, рН 7,4, 150 мМ NaCl, 1% NP-40, 0,5% дезоксихолевая кислота, 0,1% SDS) с добавлением полного коктейля ингибиторов протеазы (Sigma). После лизиса клеток буфер заменяли 0,05% TBST с использованием колонки Vivaspin (MWCO = 10 кДа) (GE Healthcare). Иммунопреципитацию проводили из 200 мкг лизатов с 6 мкг антител к CD44 (3 мкг антител Abcam к CD44 (кат. ab157107) и 3 мкг антител Proteintech к CD44 (кат. 15675-1-AP)), нормальные кроличьи IgG (Cell Signaling Technology), антитела против FLAG (M2; sigma) или нормальные мышиные IgG (CST) с использованием магнитных шариков Dynabeads Protein G (Thermo Fisher Scientific). Антитела перекрестно связывали с гранулами с помощью DMP (Thermo Fisher Scientific) перед иммунопреципитацией. Иммунопреципитанты элюировали путем инкубации с RIPA-буфером с добавлением 50 мМ ДТТ в течение 20 мин при 70°С.
ab157107) и 3 мкг антител Proteintech к CD44 (кат. 15675-1-AP)), нормальные кроличьи IgG (Cell Signaling Technology), антитела против FLAG (M2; sigma) или нормальные мышиные IgG (CST) с использованием магнитных шариков Dynabeads Protein G (Thermo Fisher Scientific). Антитела перекрестно связывали с гранулами с помощью DMP (Thermo Fisher Scientific) перед иммунопреципитацией. Иммунопреципитанты элюировали путем инкубации с RIPA-буфером с добавлением 50 мМ ДТТ в течение 20 мин при 70°С.
Масс-спектрометрия
CD44-иммунопреципитанты расщепляли трипсином и очищали с использованием колонки S-Trap (Protifi). Пептиды были отправлены в Ресурсную лабораторию масс-спектрометрии Университета Рочестера для 10-плексного мечения ТМТ и масс-спектрометрии. Образцы разделяли с помощью ионизации наноэлектрораспылением на приборе Orbitrap Fusion Lumos MS. Отнесение пептидов проводили с использованием Proteome Discoverer и Sequest, а ионы MS3 использовали для количественного определения содержания белка.
Транскриптомный анализ
РНК экстрагировали с использованием Trizol (Invitrogen). Образцы РНК были отправлены в Исследовательский центр геномики Университета Рочестера для RNA-Seq. Набор для подготовки образцов РНК TruSeq V2 (Illumina) использовали для создания библиотеки. Библиотеки гибридизовали с проточной кюветой Illumina с одним концом и амплифицировали с использованием cBot (Illumina) в концентрации 8 пМ на дорожку. Для каждого образца были сгенерированы чтения с одним концом из 100 нуклеотидов. Секвенирование проводили с использованием высокопроизводительного прибора Illumina HiSeqTM 2500. Нормализация и дифференциальный анализ экспрессии выполнялись с использованием DESeq2 46 . Анализ онтологии и путей был выполнен с использованием программы Enrichr 26 . Анализ обогащения TFBS проводили с использованием программы oPOSSUM 27 . Параметры, использованные в oPOSSUM, были следующими: отсечение консервации = 0,4, порог оценки матрицы = 85%, анализируемые последовательности = 2 kb выше сайтов начала транскрипции, отсечение по Z-оценке = 10, отсечение по шкале Фишера = 7.
Количественное определение внутриклеточных АФК
Уровни внутриклеточных АФК определяли количественно с использованием флуоресцентного зонда H 2 DCFDA. Клетки предварительно инкубировали в течение 6 часов с 20 мкг/мл NSF-HA или эквивалентным объемом PBS, затем с 5 мкМ H 2 DCFDA в HBSS в течение 30 минут, а затем подвергали воздействию 200 мкМ tBHP в течение 1 часа и промывают HBSS. Флуоресценцию DCF измеряли при возбуждении при Ex485/Em530 нм и нормализовали по количеству клеток.
Вестерн-блоттинг
Лизаты цельных клеток готовили путем лизиса клеток в лизирующем буфере (20 мМ Трис, pH 7,5, 250 мМ NaCl, 1% N-лаурилсаркозин, 10 мМ NaF, 2 мМ ортованадата натрия, 1 мМ β-глицерофосфат) с добавлением полного коктейля ингибиторов протеазы (Sigma). Ядерные и цитоплазматические лизаты готовили с использованием набора ядерных экстрактов (abcam, ab113474). Лизаты и иммунопреципитаты денатурировали при 95°С (70°С для обнаружения CD44) в течение 10 мин в буфере Лэммли. Белки разделяли с помощью SDS-PAGE, переносили на нитроцеллюлозные мембраны, блокировали 5% BSA или 5% молока в TBST в течение 1 ч и зондировали следующими первичными антителами: анти-β-актин (sc-47778; Санта-Крус), анти-CD44 (ab157107; abcam), анти-FLAG (M2; сигма), анти-GFP (для обнаружения YFP) (ab6556; abcam), анти-h4 (#9715; CST), анти-p53 (человек) (10442 -1-AP; Proteintech), анти-p53 (мышь) (ab26; Abcam), анти-фосфо-p53 (Ser-9) (#9288; CST), анти-фосфо-p53 (Ser-15) (#9286; CST), анти-фосфо-p53 (Ser-46) (#2521; CST) и анти-фосфо-p53 (Thr-81) (# 2676; КНТ). Все первичные антитела разводили до концентрации 1 мкг/мл. После промывки TBST мембраны инкубировали со вторичными антителами (Sigma) (1:1000) в течение 1 ч при комнатной температуре (КТ), снова промывали TBST и визуализировали реагентами с усилением хемилюминесценции. Необрезанные изображения геля представлены на дополнительном рисунке 10.
Белки разделяли с помощью SDS-PAGE, переносили на нитроцеллюлозные мембраны, блокировали 5% BSA или 5% молока в TBST в течение 1 ч и зондировали следующими первичными антителами: анти-β-актин (sc-47778; Санта-Крус), анти-CD44 (ab157107; abcam), анти-FLAG (M2; сигма), анти-GFP (для обнаружения YFP) (ab6556; abcam), анти-h4 (#9715; CST), анти-p53 (человек) (10442 -1-AP; Proteintech), анти-p53 (мышь) (ab26; Abcam), анти-фосфо-p53 (Ser-9) (#9288; CST), анти-фосфо-p53 (Ser-15) (#9286; CST), анти-фосфо-p53 (Ser-46) (#2521; CST) и анти-фосфо-p53 (Thr-81) (# 2676; КНТ). Все первичные антитела разводили до концентрации 1 мкг/мл. После промывки TBST мембраны инкубировали со вторичными антителами (Sigma) (1:1000) в течение 1 ч при комнатной температуре (КТ), снова промывали TBST и визуализировали реагентами с усилением хемилюминесценции. Необрезанные изображения геля представлены на дополнительном рисунке 10.
РНКи
РНКи проводили путем трансфекции киРНК с использованием реагента для трансфекции RNAiMAX (Thermo Fisher Scientific). Конечная концентрация миРНК составляла 50 нМ. Последовательности олигонуклеотидов siRNA были следующими. si-люцифераза: CGUACGCGGAAUACUUCGAtt. si-CD44: GAACGAAUCCUGAAGACAUCUtt 47 . si-RHAMM: GCUAGAUAUUGCCCAGUUAtt 48 . si-p53: AAGACUCCAGUGGUAAUCUACtt 35 .
Конечная концентрация миРНК составляла 50 нМ. Последовательности олигонуклеотидов siRNA были следующими. si-люцифераза: CGUACGCGGAAUACUUCGAtt. si-CD44: GAACGAAUCCUGAAGACAUCUtt 47 . si-RHAMM: GCUAGAUAUUGCCCAGUUAtt 48 . si-p53: AAGACUCCAGUGGUAAUCUACtt 35 .
Статистика и воспроизводимость
Столбики погрешностей представлены в виде средних значений ± стандартное отклонение. Для оценки статистической значимости использовали двусторонний критерий Стьюдента t и односторонний ANOVA с апостериорным двусторонним критерием Даннета, если не указано иное. Точные значения p указаны в файле исходных данных. Все реплики являются биологическими.
Сводка отчетов
Дополнительную информацию о дизайне исследования можно найти в Резюме отчета об исследованиях в природе, связанном с этой статьей.
Доступность данных
Исходные данные, лежащие в основе рис. 1a, b, d, e, 2b, c, e, f, i, 4b–d, f, g и дополнительные рис. 1A–C, 2A–H, 3B–E, G–I, 4A, B, 5A–C, 6B, 7E, G, 8A–F, 9B представлены в виде файла исходных данных. Данные RNA-Seq депонированы в базе данных Gene Expression Omnibus (GEO) под регистрационным номером GSE116759 [https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE116759.]. Данные масс-спектрометрии депонированы в ProteomeXchange с идентификатором PXD013445 [https://www.ebi.ac.uk/pride/archive/projects/PXD013445].
1A–C, 2A–H, 3B–E, G–I, 4A, B, 5A–C, 6B, 7E, G, 8A–F, 9B представлены в виде файла исходных данных. Данные RNA-Seq депонированы в базе данных Gene Expression Omnibus (GEO) под регистрационным номером GSE116759 [https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE116759.]. Данные масс-спектрометрии депонированы в ProteomeXchange с идентификатором PXD013445 [https://www.ebi.ac.uk/pride/archive/projects/PXD013445].
Ссылки
Edrey, Y.H., Hanes, M., Pinto, M., Mele, J. & Buffenstein, R. Успешное старение и устойчивое хорошее здоровье голого землекопа: долгоживущая модель млекопитающих для биогеронтологии и биомедицинские исследования. ИЛАР Дж. 52 , 41–53 (2011).
КАС пабмед Статья Google ученый
Руби, Дж. Г., Смит, М. и Баффенштейн, Р. Смертность голых землекопов противоречит законам Гомперца, поскольку не увеличивается с возрастом.
Elife 7 , e31157 (2018).О’Коннор, Т. П., Ли, А., Джарвис, Дж. У. и Баффенштейн, Р. Увеличение продолжительности жизни у голых землекопов: возрастные изменения метаболизма, состава тела и функции желудочно-кишечного тракта. Сравнительная биохимия и физиология. Часть А, Мол. интегр. Физиол. 133 , 835–842 (2002).
Артикул Google ученый
Буффенштейн, Р. Незначительное старение у самого долгоживущего грызуна, голого землекопа: взгляды успешно стареющего вида. Дж. Комп. Физиол. B 178 , 439–445 (2008).
ПабМед Статья Google ученый
Andziak, B. et al. Высокий уровень окислительного повреждения у самого долгоживущего грызуна — голого землекопа. Aging Cell 5 , 463–471 (2006).
КАС пабмед Статья Google ученый
Перес В.И. и др. Белковая стабильность и устойчивость к окислительному стрессу являются определяющими факторами долголетия самого долгоживущего грызуна — голого землекопа. Проц. Натл акад. науч. США 106 , 3059–3064 (2009 г.).
ОБЪЯВЛЕНИЕ КАС пабмед Статья ПабМед Центральный Google ученый
Родригес, К. А., Эдрей, Ю. Х., Осмульски, П., Гачинска, М. и Баффенштейн, Р. Измененный состав протеасомных комплексов печени способствует усилению протеасомной активности у исключительно долгоживущего голого землекопа.
PLoS ONE 7 , e35890 (2012).ОБЪЯВЛЕНИЕ КАС пабмед ПабМед Центральный Статья Google ученый
Чжао, С. и др. Высокая аутофагия у голого землекопа может играть важную роль в поддержании хорошего здоровья. Клеточная физиол. Biochem 33 , 321–332 (2014).
КАС пабмед Статья Google ученый
Чжао Ю. и др. Голые землекопы могут подвергаться клеточному старению, вызванному развитием, онкогенами и повреждением ДНК. Проц. Натл акад. науч. США 115 , 1801–1806 (2018).
КАС пабмед Статья ПабМед Центральный Google ученый
Тянь, X. и др. Локус INK4 резистентного к опухоли грызуна, голого землекопа, экспрессирует функциональную гибридную изоформу p15/p16. Проц.
Натл акад. науч. США 112 , 1053–1058 (2015).ОБЪЯВЛЕНИЕ КАС пабмед Статья Google ученый
Miyawaki, S. et al. Опухолевая резистентность индуцированных плюрипотентных стволовых клеток, полученных от голых землекопов. Нац. коммун. 7 , 11471 (2016).
ОБЪЯВЛЕНИЕ КАС пабмед ПабМед Центральный Статья Google ученый
Селуанов А. и др. Гиперчувствительность к контактному торможению дает ключ к устойчивости голого землекопа к раку. Проц. Натл акад. науч. США 106 , 19352–19357 (2009 г.).
ОБЪЯВЛЕНИЕ КАС пабмед Статья ПабМед Центральный Google ученый
Тянь, X. и др. Гиалуронан с высокой молекулярной массой опосредует устойчивость голого землекопа к раку.
Природа 499 , 346–349 (2013).ОБЪЯВЛЕНИЕ КАС пабмед ПабМед Центральный Статья Google ученый
Богумилицки Л. и др. Замаскированный обманщик: амбивалентные роли гиалуроновой кислоты в матрице. Фронт Онкол. 7 , 242 (2017).
ПабМед ПабМед Центральный Статья Google ученый
Cyphert, JM, Trempus, C.S. & Garantziotis, S. Размер имеет значение: специфичность молекулярной массы эффектов гиалуроновой кислоты в клеточной биологии. Int J. Cell Biol. 2015 , 563818 (2015).
ПабМед ПабМед Центральный Статья КАС Google ученый
Коуман, М.К., Ли, Х.Г., Швертфегер, К.Л., Маккарти, Дж.Б. и Терли, Э.А. Содержание и размер гиалуроновой кислоты в биологических жидкостях и тканях.
Перед. Иммунол. 6 , 261 (2015).ПабМед ПабМед Центральный Статья КАС Google ученый
Ян, К. и др. Высоко- и низкомолекулярные формы гиалуроновой кислоты по-разному влияют на кластеризацию CD44. Дж. Биол. хим. 287 , 43094–43107 (2012).
КАС пабмед ПабМед Центральный Статья Google ученый
Jiang, D., Liang, J. & Noble, P.W. Гиалуронан как иммунорегулятор при заболеваниях человека. Физиол. Ред. 91 , 221–264 (2011).
КАС пабмед Статья Google ученый
Pauloin, T., Dutot, M., Warnet, J.M. & Rat, P. Модуляция токсичности консервантов in vitro: гиалуронан с высокой молекулярной массой снижает апоптоз и окислительный стресс, вызванные хлоридом бензалкония.
евро. Дж. Фарм. науч. 34 , 263–273 (2008).КАС пабмед Статья Google ученый
Ye, J. et al. Цитопротекторный эффект гиалуроновой кислоты и гидроксипропилметилцеллюлозы против повреждения ДНК, вызванного тимеросалом в клетках конъюнктивы Чанга. Арка Грефеса. клин. Эксп. Офтальмол. 250 , 1459–1466 (2012).
КАС пабмед Статья Google ученый
Онодера, Ю., Терамура, Т., Такехара, Т. и Фукуда, К. Гиалуроновая кислота регулирует ключевой окислительно-восстановительный фактор Nrf2 посредством фосфорилирования Akt в бычьих суставных хондроцитах. FEBS Open Bio 5 , 476–484 (2015).
КАС пабмед ПабМед Центральный Статья Google ученый
Ву, В. и др. Защитная роль гиалуроновой кислоты при индуцированном Cr(VI) окислительном повреждении эпителиальных клеток роговицы.
J. Офтальмол. 2017 , 3678586 (2017).ПабМед ПабМед Центральный Google ученый
Laurent, U.B. & Tengblad, A. Определение гиалуроновой кислоты в биологических образцах с помощью специального метода радиоанализа. Анал. Биохим. 109 , 386–394 (1980).
КАС пабмед Статья Google ученый
Rouillard, A.D. et al. Harmonizome: коллекция обработанных наборов данных, собранных для предоставления и извлечения знаний о генах и белках. База данных (Оксфорд) 2016 (2016).
Chen, E.Y. et al. Enrichr: интерактивный и совместный инструмент анализа обогащения списка генов HTML5. БМС Биоинформа. 14 , 128 (2013).
Артикул Google ученый
Ho Sui, S.J. et al.
oPOSSUM: идентификация чрезмерно представленных сайтов связывания факторов транскрипции в коэкспрессируемых генах. Рез. нуклеиновых кислот. 33 , 3154–3164 (2005).ПабМед ПабМед Центральный Статья КАС Google ученый
Bourguignon, L.Y., Gilad, E., Rothman, K. & Peyrollier, K. Взаимодействие гиалуронана-CD44 с IQGAP1 способствует передаче сигналов Cdc42 и ERK, что приводит к связыванию актина, активации транскрипции Elk-1/рецептора эстрогена и яичникам. прогрессирование рака. Дж. Биол. хим. 280 , 11961–11972 (2005 г.).
КАС пабмед Статья Google ученый
Чжан, Л. С., Ма, Х. В., Грейнер, Х. Дж., Зуо, В. и Маммерт, М. Э. Ингибирование пролиферации клеток с помощью CD44: Akt инактивирован, а EGR-1 подавляется. Клеточный пролиф. 43 , 385–395 (2010).
КАС пабмед Статья Google ученый
Miletti-Gonzalez, K. E. et al. Идентификация функции внутрицитоплазматического домена CD44 (CD44-ICD): модуляция транскрипции матриксной металлопротеиназы 9 (MMP-9) с помощью нового ответного элемента промотора. Дж. Биол. хим. 287 , 18995–19007 (2012).
КАС пабмед ПабМед Центральный Статья Google ученый
Фишер, М. Перепись и оценка генов-мишеней p53. Онкоген 36 , 3943–3956 (2017).
КАС пабмед ПабМед Центральный Статья Google ученый
Лакнер, Д. Х., Хаяши, М. Т., Чезаре, А. Дж. и Карлседер, Дж. Геномный подход определяет регуляцию экспрессии генов, специфичную для старения. Стареющая ячейка 13 , 946–950 (2014).
КАС пабмед ПабМед Центральный Статья Google ученый
Танака Х. и др. Ген рибонуклеотидредуктазы, участвующий в p53-зависимой контрольной точке клеточного цикла для повреждения ДНК. Природа 404 , 42–49 (2000).
ОБЪЯВЛЕНИЕ КАС пабмед Статья Google ученый
Оцука Т. и др. ASC представляет собой адаптер Bax и регулирует путь митохондриального апоптоза p53-Bax. Нац. Клеточная биол. 6 , 121–128 (2004).
КАС пабмед Статья Google ученый
Керли-Гамильтон, Дж. С., Пайк, А. М., Ли, Н., Ди Ренцо, Дж. И Спинелла, М. Дж. Доминирующий p53 транскрипционный ответ на цисплатин при эмбриональной карциноме человека, происходящей из зародышевых клеток яичка. Онкоген 24 , 6090–6100 (2005).
КАС пабмед Статья Google ученый
Джекс, Т. и др. Анализ спектра опухолей у мышей с мутацией p53. Курс. биол. 4 , 1–7 (1994).
КАС пабмед Статья Google ученый
Fischer, M. Сохранение и расхождение регуляторной сети гена p53 между мышами и людьми. Онкоген 38 , 4095–4109 (2019).
КАС пабмед ПабМед Центральный Статья Google ученый
Кин, М. и др. Ресурс генома голых землекопов: содействие анализу рака и адаптации, связанной с долголетием. Биоинформатика 30 , 3558–3560 (2014).
КАС пабмед ПабМед Центральный Статья Google ученый
Suwan, K. et al. Versican/PG-M собирает гиалуронан во внеклеточный матрикс и ингибирует CD44-опосредованную передачу сигналов в направлении преждевременного старения в эмбриональных фибробластах.
Дж. Биол. хим. 284 , 8596–8604 (2009 г.).КАС пабмед ПабМед Центральный Статья Google ученый
Куо, Х.Л., Хо, П.С., Хуанг, С.С. и Чанг, Н.С. В погоне за передачей сигналов с помощью трехмолекулярной покадровой FRET-микроскопии. Дисков клеточной смерти. 4 , 45 (2018).
ПабМед ПабМед Центральный Статья КАС Google ученый
Милваганам, С. М., Гринштейн, С. и Фриман, С. А. Пикеты в плазматической мембране: функции в иммунных клетках и фагоцитоз. Семин Иммунопатол. 40 , 605–615 (2018).
КАС пабмед Статья Google ученый
Уэрта-Анжелес Г., Неспорова К., Амброзова Г., Кубала Л. и Велебны В. Эффективный перевод: разработка медицинских препаратов на основе гиалуроновой кислоты с физико-химических и доклинических аспектов.
Перед. биоинж. Биотехнолог. 6 , 62 (2018).ПабМед ПабМед Центральный Статья Google ученый
Селуанов А., Вайдя А., Горбунова В. Получение первичных культур взрослых фибробластов грызунов. Дж. Виз. Эксп. 44 , 2033 (2010).
Донг Ю. и др. Не содержащие эндотоксинов гиалуронан и гиалуронановые фрагменты не стимулируют ФНО-альфа, интерлейкин-12 и не активируют костимулирующие молекулы в дендритных клетках или макрофагах. Науч. Респ. 6 , 36928 (2016).
ОБЪЯВЛЕНИЕ КАС пабмед ПабМед Центральный Статья Google ученый
Биттер, Т. и Мьюир, Х.М. Модифицированная реакция карбазола уроновой кислоты. Анал. Биохим. 4 , 330–334 (1962).
КАС пабмед Статья Google ученый
Palyi-Krekk, Z. et al. Индуцированная гиалуроновой кислотой маскировка ErbB2 и усиленная CD44 интернализация трастузумаба при резистентном к трастузумабу раке молочной железы. евро. Дж. Рак 43 , 2423–2433 (2007).
КАС пабмед Статья Google ученый
Gao, C.L.S., Wang, Y., Cha, G. & Xu, X. Влияние рецептора ингибирования подвижности, опосредованного гиалуроновой кислотой, на радиочувствительность аденокарциномы легкого A549клетки. Перевод. Рак Рез. 8 , 410–421 (2019).
КАС Статья пабмед ПабМед Центральный Google ученый
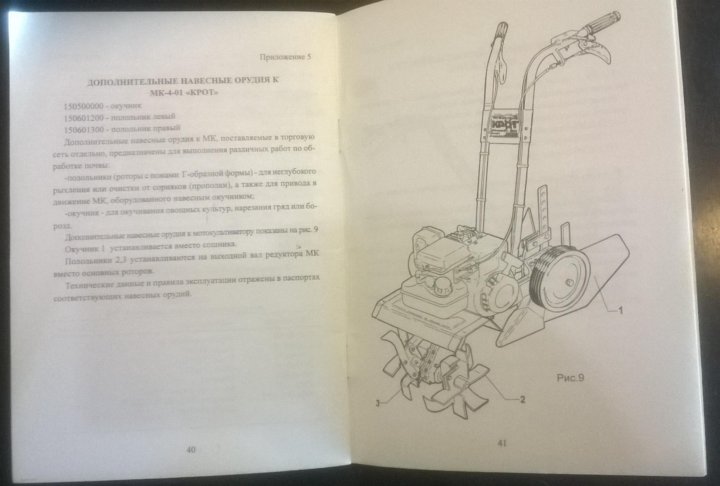 Elife 7 , e31157 (2018).
Elife 7 , e31157 (2018). «>
«>Де Ваал, Э. М. и др. Повышенное карбонилирование белка и окислительный стресс не влияют на структуру и функцию белка у долгоживущих голых землекопов: протеомный подход. Биохим. Биофиз. Рез. коммун. 434 , 815–819 (2013).
ПабМед Статья КАС Google ученый
 PLoS ONE 7 , e35890 (2012).
PLoS ONE 7 , e35890 (2012). Натл акад. науч. США 112 , 1053–1058 (2015).
Натл акад. науч. США 112 , 1053–1058 (2015). Природа 499 , 346–349 (2013).
Природа 499 , 346–349 (2013). Перед. Иммунол. 6 , 261 (2015).
Перед. Иммунол. 6 , 261 (2015). евро. Дж. Фарм. науч. 34 , 263–273 (2008).
евро. Дж. Фарм. науч. 34 , 263–273 (2008). J. Офтальмол. 2017 , 3678586 (2017).
J. Офтальмол. 2017 , 3678586 (2017). oPOSSUM: идентификация чрезмерно представленных сайтов связывания факторов транскрипции в коэкспрессируемых генах. Рез. нуклеиновых кислот. 33 , 3154–3164 (2005).
oPOSSUM: идентификация чрезмерно представленных сайтов связывания факторов транскрипции в коэкспрессируемых генах. Рез. нуклеиновых кислот. 33 , 3154–3164 (2005).


 Дж. Биол. хим. 284 , 8596–8604 (2009 г.).
Дж. Биол. хим. 284 , 8596–8604 (2009 г.). Перед. биоинж. Биотехнолог. 6 , 62 (2018).
Перед. биоинж. Биотехнолог. 6 , 62 (2018). «>
«>Лав, М. И., Хубер, В. и Андерс, С. Модерированная оценка изменения кратности и дисперсии для данных секвенирования РНК с помощью DESeq2. Геном Биол. 15 , 550 (2014).
ПабМед ПабМед Центральный Google ученый
Скачать ссылки
Благодарности
Эта работа была поддержана грантом Национального института здравоохранения США (NIH) В.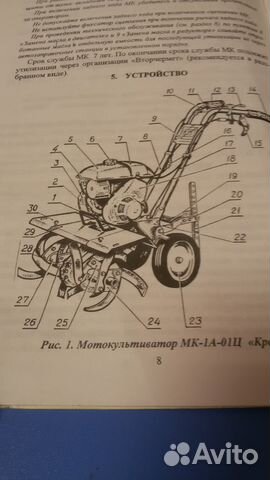 Г. и в качестве. М.Т. был частично поддержан Японским обществом содействия развитию науки (JSPS) и Мемориальным фондом Уэхара.
Г. и в качестве. М.Т. был частично поддержан Японским обществом содействия развитию науки (JSPS) и Мемориальным фондом Уэхара.
Информация о авторе
Авторы и принадлежности
Департамент биологии, Университет Рочестера, Рочестер, Нью -Йорк, 14627, США
Масаки Такасуги, Денис Фирсанов, Грегори Томблин, Юлия Аблава, Андрея -Сель -Селаун. Отделение болезней органов пищеварения, Первая дочерняя больница Университета Чжэнчжоу, Чжэнчжоу, Чжэнчжоу, Хэнань, 450052, Китайская Народная Республика
Ханьбин Нин
Авторы
- Масаки Такасуги
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Денис Фирсанов
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Gregory Tombline
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Академия
- Hanbing Ning
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Юлия Аблаева
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Андрей Селуанов
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Вера Горбунова
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Вклады
М. Т., А.С. и В.Г. задумал и спроектировал исследование. М.Т., Д.Ф., Г.Т., Х.Н. и Дж.А. проводил опыты. М.Т., А.С., В.Г. написал рукопись.
Т., А.С. и В.Г. задумал и спроектировал исследование. М.Т., Д.Ф., Г.Т., Х.Н. и Дж.А. проводил опыты. М.Т., А.С., В.Г. написал рукопись.
Авторы переписки
Переписка с Андрей Селуанов или Вера Горбунова.
Декларации этики
Конкурирующие интересы
Авторы заявляют об отсутствии конкурирующих интересов.
Дополнительная информация
Информация о рецензировании Nature Communications благодарит Robert Shmookler Reis, Spyridon Skandalis и других анонимных рецензентов за их вклад в рецензирование этой работы. Доступны отчеты о рецензировании.
Примечание издателя Springer Nature остается нейтральной в отношении юрисдикционных претензий в опубликованных картах и институциональной принадлежности.
Supplementary information
Supplementary Information
Peer Review File
Supplementary Data1
Reporting Summary
source data
Source Data
Rights and permissions
Open Access Эта статья находится под лицензией Creative Commons Attribution 4. 0 International License, которая разрешает использование, совместное использование, адаптацию, распространение и воспроизведение на любом носителе или в любом формате при условии, что вы укажете первоначальных авторов и источник, а также ссылку на лицензию Creative Commons и указать, были ли внесены изменения. Изображения или другие сторонние материалы в этой статье включены в лицензию Creative Commons для статьи, если иное не указано в кредитной строке материала. Если материал не включен в лицензию Creative Commons статьи, а ваше предполагаемое использование не разрешено законом или выходит за рамки разрешенного использования, вам необходимо получить разрешение непосредственно от правообладателя. Чтобы просмотреть копию этой лицензии, посетите http://creativecommons.org/licenses/by/4.0/.
0 International License, которая разрешает использование, совместное использование, адаптацию, распространение и воспроизведение на любом носителе или в любом формате при условии, что вы укажете первоначальных авторов и источник, а также ссылку на лицензию Creative Commons и указать, были ли внесены изменения. Изображения или другие сторонние материалы в этой статье включены в лицензию Creative Commons для статьи, если иное не указано в кредитной строке материала. Если материал не включен в лицензию Creative Commons статьи, а ваше предполагаемое использование не разрешено законом или выходит за рамки разрешенного использования, вам необходимо получить разрешение непосредственно от правообладателя. Чтобы просмотреть копию этой лицензии, посетите http://creativecommons.org/licenses/by/4.0/.
Перепечатка и разрешения
Об этой статье
Дополнительная литература
Содержание и размер гиалуроновой кислоты в тканях и плазме голого землекопа
- Дельфин дель Мармоль
- Сюзанна Хольце
- Бруно Фламион
Научные отчеты (2021)
Комментарии
Отправляя комментарий, вы соглашаетесь соблюдать наши Условия и Правила сообщества. Если вы обнаружите что-то оскорбительное или не соответствующее нашим условиям или правилам, отметьте это как неприемлемое.
Если вы обнаружите что-то оскорбительное или не соответствующее нашим условиям или правилам, отметьте это как неприемлемое.
Борьба с ущербом от кротов и полевок
5 минут чтения
Повреждение вашего имущества может быть вызвано кротами или полевками. Знание разницы между двумя животными необходимо для решения проблемы.
Кроты и полевки, хотя и прокладывают туннели, различаются по поведению и типу вреда, который они наносят газонам, садам и сельскому хозяйству. Из-за этого методы, используемые для контроля их деятельности, неодинаковы. Прежде чем действовать, вы должны знать, кто ваш противник.
Рис. 1. Крот (а) связан с землеройкой. У него длинный нос и перепончатые передние лапы с острыми когтями для копания. Полевка (b) из семейства грызунов меньше по размеру, у нее короткий хвост и ноги.
Кроты и полевки физически разные.
Южные кроты ( Scalopus aquaticus) имеют длину от 6 до 7 дюймов и вес от 3 до 4 унций. У них маленькие глазные и ушные отверстия и заостренный нос, который выступает примерно на полдюйма за пределы рта. Их большие передние лапы перепончатые и имеют острые когти, помогающие копать землю (рис. 1а).
У них маленькие глазные и ушные отверстия и заостренный нос, который выступает примерно на полдюйма за пределы рта. Их большие передние лапы перепончатые и имеют острые когти, помогающие копать землю (рис. 1а).
Полевки имеют длину от 4 до 6 дюймов, короткие ноги и хвост, а также маленькие глаза и уши. В Алабаме обитают два вида. Наиболее распространена сосновая или лесная полевка ( Microtus pinetorum), которая встречается в лесах по всему штату (рис. 1b). Вторая — луговая полевка ( Microtus ochrogaster), встречающаяся только в северной трети штата.
Кроты — хищники; полевки травоядные.
Кроты питаются белыми личинками, дождевыми червями, жуками и различными личинками. На самом деле они могут приносить пользу растениям, питаясь личинками и червями, которые повреждают растения. Полевки питаются травой, разнотравьем и иногда корой деревьев.
У кротов и полевок разные места обитания.
Кроты делают знакомую систему приподнятых туннелей на газонах. Обычно они живут поодиночке, хотя самки и детеныши могут жить в одной норе. Они строят кормовые туннели и гнездовые норы в сухой, более теплой почве под деревьями или твердыми конструкциями. Кажется, они предпочитают прохладную влажную почву (то же самое предпочитают личинки и дождевые черви).
Обычно они живут поодиночке, хотя самки и детеныши могут жить в одной норе. Они строят кормовые туннели и гнездовые норы в сухой, более теплой почве под деревьями или твердыми конструкциями. Кажется, они предпочитают прохладную влажную почву (то же самое предпочитают личинки и дождевые черви).
У полевок есть подземные туннели. Они ищут пищу на домашнем участке площадью около четверти акра и редко выходят на открытые места. Их предпочтительная среда обитания — районы с густым напочвенным покровом, такие как естественная среда, фруктовые сады, поля и сады.
Полевки производят больше потомства.
Полевки размножаются круглый год. У них может быть до пяти пометов из шести потомков. Продолжительность жизни полевки составляет всего от 2 до 16 месяцев. Популяции имеют тенденцию достигать пика каждые 2–5 лет. В это время плотность полевок может стать довольно высокой, а участки, на которых не было повреждений, могут внезапно получить серьезные повреждения. Кроты размножаются в марте и апреле. Беременность составляет примерно 5 недель, а размер помета колеблется от двух до пяти потомков. Из-за одиночного характера кротов вы можете найти только пять или шесть кротов на акр.
Беременность составляет примерно 5 недель, а размер помета колеблется от двух до пяти потомков. Из-за одиночного характера кротов вы можете найти только пять или шесть кротов на акр.
Полевки убивают растения; родинок обычно нет.
Люди, которые пострадали от полевки, обычно описывают такие сценарии: Однажды мое маленькое дерево выглядело здоровым, а на следующий день оно погибло. Или: Однажды мой цветник был прекрасен, а на следующий день растения завяли и погибли. При ближайшем рассмотрении можно обнаружить крошечные следы зубов вокруг растения на уровне земли или отсутствие корневой системы.
Кроты редко причиняют значительный вред растениям. Однако их деятельность по прокладыванию туннелей может обезображивать газоны и сады (рис. 2 и рис. 3). Наибольший риск представляют норы, которые размываются во время проливных дождей, что представляет угрозу безопасности. Наиболее очевидным признаком повреждения полевками является мертвое или отмирающее растение. Восприимчивы огороды, декоративные насаждения и молодняк леса. Полевки могут туннелировать рядом с корневой системой, поедая корни и пережевывая или «опоясывая» основной стебель прямо над землей.
Полевки могут туннелировать рядом с корневой системой, поедая корни и пережевывая или «опоясывая» основной стебель прямо над землей.
- Рисунок 2. Промытая нора кротов может обезобразить газоны.
- Рисунок 3. Эта линия голой земли — место, где крот прорыл туннель через двор.
- Рис. 4. Ствол небольшого кизила со всей корневой структурой, изъеденной полёвками. Домовладелец сообщил, что однажды с деревом все было в порядке; на следующий день листья завяли. А на следующий день домовладелец без особых усилий вытащил дерево из земли, и это все, что от него осталось.
Повреждение полёвками можно спутать с повреждением кроликами. Чтобы определить виновника, посмотрите на грызущий или жевательный рисунок. У полевок маленькие зубы, которые оставляют на растении маленькие неравномерные следы от выгрызаний под разными углами (рис. 4). У кроликов более широкие зубы, которые оставляют более широкие следы. Кроме того, кролики часто разрезают растение пополам с одинаковым срезом под углом 45 градусов.
Кроме того, кролики часто разрезают растение пополам с одинаковым срезом под углом 45 градусов.
Репелленты и ядовитые вещества, как правило, неэффективны для борьбы с повреждением кротов. Одна из трудностей с ядовитыми веществами заключается в том, чтобы заставить кротов принять приманку. Меры летального или биологического контроля являются наиболее эффективными.
Ловушки
Смертельные ловушки обычно бывают трех типов: гарпунные (рис. 6), ножницы-челюсти (рис. 8) и колье. Любой из них работает хорошо, если настроен правильно, но тип почвы может повлиять на эффективность. На песчаных почвах более эффективны гарпунные ловушки, на суглинистых почвах или почвах с более высоким содержанием глины более эффективны ловушки с ножницами и чокерами.
Прежде чем устанавливать ловушки, вы должны определить, какие ходы или ходы для кротов используются чаще всего. Чтобы выяснить это, выровняйте туннели, наступив на них или используя газонный каток. На следующий день посмотрите, какие из них выскочили обратно. Ставьте ловушки в эти новые туннели.
Ставьте ловушки в эти новые туннели.
Если вы используете гарпунную ловушку, дайте ей несколько раз прыгнуть в землю, прежде чем сделать последний бросок. Это гарантирует, что гарпуны могут беспрепятственно пройти через туннель.
- Рисунок 5. Гарпунные ловушки (слева) эффективны в песчаных почвах, а ловушки с ножницами (справа) эффективны в суглинистых почвах.
- Рисунок 6. Гарпунная ловушка установлена и готова к размещению в туннеле для кротов.
- Рисунок 7. При установке ловушки с ножницами выкопайте часть почвы вокруг туннеля.
- Рисунок 8. Эта ловушка с ножницами установлена и готова к размещению в тоннеле для кротов.
- Рисунок 9. Расположите ловушку так, чтобы челюсти прорезали туннель и убили крот.
- Рисунок 10. Используйте выкопанную почву или листья, чтобы заполнить участки ямы. Главное, чтобы в нору не попадал солнечный свет.

При установке ловушек с ножницами или кольцеобразных ловушек выкопайте часть почвы вокруг туннеля и поместите ловушку в отверстие (рис. 7, 8 и 9). Снова засыпьте яму землей (рис. 10), следя за тем, чтобы свет не проникал в туннель. Рекомендуется надевать резиновые или латексные перчатки, чтобы ваш запах не попал на ловушку. После установки ловушки следите за тем, чтобы не ходить и не тревожить другие части туннелей.
Иногда ловушки могут срабатывать без поимки крота, поэтому ежедневно проверяйте ловушки и при необходимости перезагружайте их. Если крот не использует туннель с ловушкой через несколько дней, переместите ловушку в другой туннель. Как только ловушка будет на месте, обязательно снимите предохранитель.
Биологический контроль
Чтобы добиться несмертельного контроля над кротами, вы должны устранить источник пищи. Это включает в себя применение инсектицидов для борьбы с популяциями личинок. С белыми личинками можно бороться естественным образом, занося в почву молочную споровую болезнь. Хотя эти методы могут быть эффективными, они не быстры. Может пройти некоторое время, прежде чем запасы пищи сократятся настолько, что это повлияет на популяцию кротов.
Хотя эти методы могут быть эффективными, они не быстры. Может пройти некоторое время, прежде чем запасы пищи сократятся настолько, что это повлияет на популяцию кротов.
Полевки обычно не выходят на открытую территорию; поэтому изменение среды обитания путем уничтожения напочвенного покрова может быть эффективным средством уменьшения ущерба.
Ловушки
Отлов полевок в крупномасштабных операциях экономически невыгоден, но может быть полезен в цветниках или небольших огородах. Ставьте ловушки размером с мышь на входе в туннели/взлётно-посадочные полосы. Насадите ловушки смесью арахисового масла и овсянки или нарезанными яблоками. Ставьте ловушки так, чтобы курок был обращен к выходу из туннеля.
Биологические средства защиты
Держите газоны, прилегающие к цветникам, подстриженными на небольшую высоту, чтобы полевки не двигались в сады для кормления. Также уменьшите количество мульчи в цветниках и часто переворачивайте мульчу, чтобы удержать их от создания туннельных систем. Очистите задние кольца мульчи или насыпи на расстоянии не менее 3 футов от основания деревьев. В сельскохозяйственных условиях обработка почвы разрушает туннельные системы. Это помогает уменьшить популяцию полевок и последующий ущерб.
Очистите задние кольца мульчи или насыпи на расстоянии не менее 3 футов от основания деревьев. В сельскохозяйственных условиях обработка почвы разрушает туннельные системы. Это помогает уменьшить популяцию полевок и последующий ущерб.
Змеи, ястребы, совы и другие хищники питаются полевками, если им предоставляется возможность. Однако полевки обладают чрезвычайно высоким репродуктивным потенциалом, поэтому сомнительно, что хищники в одиночку могут предотвратить ущерб.
Несмотря на то, что кроты могут быть полезными в борьбе с газонными насекомыми, многие люди считают их разрушительными для ландшафта и хотят их удалить. Отлов и биологический контроль — два наиболее многообещающих метода борьбы с повреждениями кротов.
Ущерб, нанесенный полёвками, может различаться по степени тяжести из года в год по мере увеличения и уменьшения численности популяции. Вред, наносимый полевками декоративным и огородным садам, может потребовать контроля путем изменения среды обитания и отлова. Комбинация методов контроля обычно дает наилучшие результаты контроля.
Комбинация методов контроля обычно дает наилучшие результаты контроля.
Загрузите документ в формате PDF «Контроль ущерба от кротов и полевок», ANR-2412.
Вы нашли это полезным?
Пространственное обучение и память у голого слепыша: эволюционная адаптация к подземной нише
Введение
Сложные системы туннелей, поддерживаемые батиергидами, свидетельствуют об продвинутых навыках пространственной навигации. Это подтверждается исследованиями, проведенными на других подземных видах (Kimchi and Terkel, 2001, 2004; Costanzo et al., 2009).; Oosthuizen и др., 2013; Брачетта и др., 2014 г.; Oosthuizen, 2020), в том числе одно исследование, которое показало, что дамараландские землекопы превосходят полевок и крыс в тестах на долговременную память (Kimchi and Terkel, 2001). Тем не менее, ограниченные исследования были сосредоточены на оптимизации лабиринтов для проверки пространственного обучения и памяти у подземных видов, и на сегодняшний день такие исследования на голых землекопах не проводились.
Голые слепыши — вид грызунов, обитающих в Восточной Африке (Sherman et al., 1991). Как и другие батиергиды, это подземные грызуны, которые процветают в жарких и влажных средах и строят сложные норы из соединенных между собой подземных туннелей (Sherman et al., 1991). Более обширные системы нор связаны с большей производительностью пространственной памяти у других видов землекопов, вероятно, из-за ежедневной зависимости от пространственных сигналов, необходимых для навигации по этим подземным лабиринтам (Costanzo et al., 2009). Эти системы включают в себя специально отведенные жилые помещения, состоящие из туалетных камер, пищевых камер и гнездовых камер (Faulkes and Bennett, 2021; Smith and Buffenstein, 2021), сложность которых также может требовать хорошо развитой пространственной памяти. Голые землекопы прокладывают системы подземных туннелей, длина которых может достигать нескольких километров (Бретт, 19 лет).91), что может быть отчасти связано с большими размерами их колоний, насчитывающими примерно до 300 особей (Faulkes and Bennett, 2021). И голые землекопы, и дамаралендские землекопы демонстрируют социальную иерархию, характеризующуюся установленным разделением труда, а также наличием одной королевы в качестве единственной репродуктивно активной самки (Wilson, 1971; Buffenstein et al., 2022). У дамаралендских слепышей большие размеры колоний способствовали улучшению навыков пространственной навигации (du Toit et al., 2012), но это еще предстоит экспериментально исследовать на голых землекопах.
И голые землекопы, и дамаралендские землекопы демонстрируют социальную иерархию, характеризующуюся установленным разделением труда, а также наличием одной королевы в качестве единственной репродуктивно активной самки (Wilson, 1971; Buffenstein et al., 2022). У дамаралендских слепышей большие размеры колоний способствовали улучшению навыков пространственной навигации (du Toit et al., 2012), но это еще предстоит экспериментально исследовать на голых землекопах.
Подземная среда обитания Bathyergidae привела к эволюционной адаптации сенсорных способностей и соответствующих анатомических и нейроанатомических систем, которые необходимо учитывать при разработке и интерпретации поведенческих тестов. Традиционно поведенческие парадигмы в лабораторных условиях разрабатывались для более типичных моделей животных (например, мышей и крыс) и включали аспекты, соответствующие их зрительным и слуховым возможностям. Эти конструкции часто не применимы к более нетипичным исследовательским моделям, таким как подземный голый землекоп (9). 0023 Heterocephalus glaber ), для которых сенсорные ограничения, такие как функциональная слепота, играют роль (Hetling et al., 2005). В частности, парадигмы тестирования пространственной памяти, такие как водный лабиринт Морриса (Morris, 1984) и лабиринт с радиальными рукавами (Olton and Samuelson, 1976; Brown and Cook, 1986), часто полагаются на визуальные подсказки, которые обеспечивают основу для навигации в лабиринте. .
0023 Heterocephalus glaber ), для которых сенсорные ограничения, такие как функциональная слепота, играют роль (Hetling et al., 2005). В частности, парадигмы тестирования пространственной памяти, такие как водный лабиринт Морриса (Morris, 1984) и лабиринт с радиальными рукавами (Olton and Samuelson, 1976; Brown and Cook, 1986), часто полагаются на визуальные подсказки, которые обеспечивают основу для навигации в лабиринте. .
Разработка поведенческих задач для подземных грызунов требует учета их сенсорных способностей, отличных от других представителей Rodentia, а также включения соответствующих мотиваторов. В текущем исследовании мы разработали эффективную парадигму для изучения пространственного обучения и памяти у голых землекопов с учетом ряда факторов, которые могут повлиять на способности навигационной памяти среди членов колонии, включая возраст, пол, массу тела и ранг доминирования в пределах социальная иерархия. Мы предположили, что голые землекопы будут демонстрировать пространственное обучение в лабиринте с индивидуальной конструкцией за счет более быстрого достижения наградной камеры (уменьшенная задержка) и с оптимизированной навигацией (более короткое общее пройденное расстояние и меньшее количество совершенных ошибок) во время тренировочных испытаний, и что пространственная память будет лучше. надежный, основанный на адаптации голого землекопа к подземной среде обитания.
надежный, основанный на адаптации голого землекопа к подземной среде обитания.
Материалы и методы
Животные
В этом исследовании использовали сорок три голых землекопа ( H. glaber; Rüppell, 1842) (25 самок, 18 самцов) различной массы тела и возраста. Для предварительного исследования с использованием конструкции прерывистого лабиринта использовали восемь животных (таблица 1). После усовершенствования конструкции лабиринта и включения непрерывного лабиринта было использовано тридцать пять животных. Шесть из этих животных были исключены из-за отсутствия данных для ранжирования их доминирования. У одиннадцати дополнительных животных отсутствовало не более одной контрольной точки данных. Для этих одиннадцати животных отсутствующая точка данных была заменена линейной интерполяцией. Одно дополнительное животное было исключено как выброс из-за того, что точки данных в первом обучающем испытании превышали десять стандартных отклонений от среднего, в то время как другое животное было исключено из-за смерти до окончания сбора данных. Таким образом, в исследование непрерывного лабиринта было включено 27 животных (табл. 1). Для анализа пробы памяти были исключены два дополнительных животных, одно как выброс со значениями, превышающими 10 стандартных отклонений от среднего, а другое исключено из-за отсутствия нескольких точек данных. Таким образом, 25 животных из исходных 27 были включены в анализ зонда памяти. Масса тела животных, включенных в анализ, варьировала от 23,0 до 101,0 г (компактная портативная масштабная модель Braintree Scientific CB 1001 с точностью до 0,1 г), возраст варьировал от 0,39до 6,68 лет в начале испытаний. Все голые землекопы были членами одной из двух лабораторных племенных колоний (колонии А и колонии В), которые содержались в доме, с разным статусом внутри колонии (королевы, производители, не размножающиеся). Не все члены каждой колонии были использованы. Животные были исключены на основании участия в других текущих исследованиях. В помещении вивария для каждой колонии поддерживалась температура приблизительно 27,8–30°C и относительная влажность не менее 40 %, без смены дня и ночи из-за подземной среды обитания животных (Sherman et al.
Таким образом, в исследование непрерывного лабиринта было включено 27 животных (табл. 1). Для анализа пробы памяти были исключены два дополнительных животных, одно как выброс со значениями, превышающими 10 стандартных отклонений от среднего, а другое исключено из-за отсутствия нескольких точек данных. Таким образом, 25 животных из исходных 27 были включены в анализ зонда памяти. Масса тела животных, включенных в анализ, варьировала от 23,0 до 101,0 г (компактная портативная масштабная модель Braintree Scientific CB 1001 с точностью до 0,1 г), возраст варьировал от 0,39до 6,68 лет в начале испытаний. Все голые землекопы были членами одной из двух лабораторных племенных колоний (колонии А и колонии В), которые содержались в доме, с разным статусом внутри колонии (королевы, производители, не размножающиеся). Не все члены каждой колонии были использованы. Животные были исключены на основании участия в других текущих исследованиях. В помещении вивария для каждой колонии поддерживалась температура приблизительно 27,8–30°C и относительная влажность не менее 40 %, без смены дня и ночи из-за подземной среды обитания животных (Sherman et al. , 19).91). Жилье для животных состояло из ряда камер, соединенных между собой трубками и спроектированных так, чтобы имитировать их естественную систему нор. Был обеспечен свободный доступ к еде и большому количеству постельных принадлежностей (дополнительную информацию о размещении см. в Artwohl et al., 2002). Все аспекты исследования, проведенного для этого исследования, соответствовали нашему протоколу, одобренному Институциональным комитетом по уходу и использованию животных Университета Южного Иллинойса, Карбондейл, Иллинойс, США, и соответствовали Руководству Национального института здравоохранения по уходу и использованию. лабораторных животных (публикации NIH № 8023 и 1978).
, 19).91). Жилье для животных состояло из ряда камер, соединенных между собой трубками и спроектированных так, чтобы имитировать их естественную систему нор. Был обеспечен свободный доступ к еде и большому количеству постельных принадлежностей (дополнительную информацию о размещении см. в Artwohl et al., 2002). Все аспекты исследования, проведенного для этого исследования, соответствовали нашему протоколу, одобренному Институциональным комитетом по уходу и использованию животных Университета Южного Иллинойса, Карбондейл, Иллинойс, США, и соответствовали Руководству Национального института здравоохранения по уходу и использованию. лабораторных животных (публикации NIH № 8023 и 1978).
Таблица 1. Описательная статистика (приведенные значения представляют собой среднее значение ± стандартная ошибка среднего и диапазон) для голых землекопов, использованных в двух различных конструкциях пространственных лабиринтов.
Дизайн лабиринта
Мы разработали модифицированные версии лабиринта Хебба-Уильямса, распространенной парадигмы для тестирования пространственного обучения и памяти у грызунов. Модификации лабиринта Хебба-Уильямса были основаны на исследованиях, в которых использовались аналогичные тесты пространственной навигации на других нетипичных животных моделях грызунов, включая подземных туко-туко (Antinuchi and Schleich, 2003; Schleich, 2010; Brachetta et al., 2014) и слепыш слепыш (Кимчи, Теркель, 2001, 2004). Лабиринты были построены с использованием труб из ПВХ, которые были разрезаны пополам в продольном направлении и покрыты листом прозрачного акрила, создавая желоб, по которому животные могли перемещаться, и обеспечивая видимость для видеозаписи. Трубка имела диаметр 2 дюйма, такой же, как и трубка в туннельных системах жилищных камер для голых землекопов. Эта ширина трубок была выбрана на основе предыдущих исследований, которые показали более высокую эффективность обучения и памяти у слепышей при использовании более узких путей, которые обеспечивают тактильные сигналы в лабиринтах (Kimchi and Terkel, 2004).
Модификации лабиринта Хебба-Уильямса были основаны на исследованиях, в которых использовались аналогичные тесты пространственной навигации на других нетипичных животных моделях грызунов, включая подземных туко-туко (Antinuchi and Schleich, 2003; Schleich, 2010; Brachetta et al., 2014) и слепыш слепыш (Кимчи, Теркель, 2001, 2004). Лабиринты были построены с использованием труб из ПВХ, которые были разрезаны пополам в продольном направлении и покрыты листом прозрачного акрила, создавая желоб, по которому животные могли перемещаться, и обеспечивая видимость для видеозаписи. Трубка имела диаметр 2 дюйма, такой же, как и трубка в туннельных системах жилищных камер для голых землекопов. Эта ширина трубок была выбрана на основе предыдущих исследований, которые показали более высокую эффективность обучения и памяти у слепышей при использовании более узких путей, которые обеспечивают тактильные сигналы в лабиринтах (Kimchi and Terkel, 2004).
Первый испытанный нами лабиринт был непрерывным и включал тупики (рис. 1А), аналогичные другим лабиринтам на основе Хебба-Уильямса (Kimchi and Terkel, 2001, 2004; Costanzo et al., 2009; du Toit et al. ., 2012). Для этого варианта лабиринта было проведено предварительное тестирование на когорте из восьми животных (табл. 1). Все тупики внутри лабиринта были запечатаны, а поверх лабиринта был помещен прозрачный акриловый лист, позволяющий визуализировать поведение и поведение животных в лабиринтах сверху. Камера вознаграждения состояла из цилиндрического корпуса из прозрачного литого акрила диаметром 8 дюймов, высотой 10 дюймов и толщиной стенки 1/4 дюйма. Эта наградная камера обогревалась тепловой лампой и заполнялась подстилкой из домашней колонии животного. Он был связан с лабиринтом через соединительную трубку из прозрачного литого акрила диаметром 2 дюйма с толщиной стенки 1/4 дюйма (US Plastics, артикул № 44530). Производительность измерялась с использованием задержки до камеры вознаграждения, общего расстояния, пройденного до достижения камеры вознаграждения, и количества входов в зоны ошибок.
1А), аналогичные другим лабиринтам на основе Хебба-Уильямса (Kimchi and Terkel, 2001, 2004; Costanzo et al., 2009; du Toit et al. ., 2012). Для этого варианта лабиринта было проведено предварительное тестирование на когорте из восьми животных (табл. 1). Все тупики внутри лабиринта были запечатаны, а поверх лабиринта был помещен прозрачный акриловый лист, позволяющий визуализировать поведение и поведение животных в лабиринтах сверху. Камера вознаграждения состояла из цилиндрического корпуса из прозрачного литого акрила диаметром 8 дюймов, высотой 10 дюймов и толщиной стенки 1/4 дюйма. Эта наградная камера обогревалась тепловой лампой и заполнялась подстилкой из домашней колонии животного. Он был связан с лабиринтом через соединительную трубку из прозрачного литого акрила диаметром 2 дюйма с толщиной стенки 1/4 дюйма (US Plastics, артикул № 44530). Производительность измерялась с использованием задержки до камеры вознаграждения, общего расстояния, пройденного до достижения камеры вознаграждения, и количества входов в зоны ошибок. Заход в зону ошибки засчитывался, если более 50 % тела животного проходило в зону, независимо от направления входа.
Заход в зону ошибки засчитывался, если более 50 % тела животного проходило в зону, независимо от направления входа.
Рис. 1. Конфигурации для тестирования поведения голого землекопа. (A) Модифицированный лабиринт Хебба-Уильямса с прерывистой конструкцией лабиринта, включающей тупики. Серые области обозначают области, в которых при входе была зафиксирована ошибка. (B) Модифицированный лабиринт Хебба-Уильямса с непрерывной конструкцией, лишенный тупиков и имеющий больше сходства с обширными взаимосвязанными системами подземных туннелей в среде обитания голого землекопа. (C) Тестовая установка мотиватора. Животных помещали в стартовую камеру и давали 2 мин для свободного исследования. Производительность животных тестировалась с использованием ряда мотиваторов в камере вознаграждения и сравнивалась с производительностью, когда эта камера была пуста. (D) Настройка тестирования доминирования. Животных помещали в указанные точки входа в каждую камеру, лицом друг к другу, и анализировали, сколько раз одно проходило над другим через трубку в трех испытаниях.
Голые землекопы сильно реагируют на засоры в туннелях, упорно копая и кусая в попытке преодолеть завал (Sherman et al., 1991). Во время первоначального тестирования дизайна лабиринта (рис. 1А) мы обнаружили, что это отвлечение влияет на завершение лабиринта («Частота неудач», таблица 1). Частота неудач рассчитывалась как процент обучающих попыток (испытания 1–15), в которых животные не прошли лабиринт за отведенные 5 минут (таблица 1).
FailRate(%)=IncompleteTrials/TotalTrials× 100(1)
Чтобы решить эту проблему, мы преобразовали лабиринт в непрерывную конфигурацию без тупиков (рис. 1B) и протестировали тридцать пять животных в этой конфигурации лабиринта (таблица 1). . Это позволило нам более эффективно исследовать, будут ли животные более мотивированы на прохождение лабиринта, не отвлекая внимание от поиска камеры с вознаграждением из-за тупиковых отвлекающих факторов.
Тестирование вознаграждения
Чтобы проверить эффективность ряда мотиваторов на голых землекопах и впоследствии использовать наиболее эффективный мотиватор в камере вознаграждения, перед использованием непрерывного лабиринта был проведен отдельный эксперимент. Те же самые восемь животных, которые использовались при тестировании первой конструкции лабиринта, использовались для определения подходящих мотиваторов для голых землекопов. Сначала в качестве контроля стартовую камеру соединяли с отдельной пустой камерой с помощью Г-образной соединительной трубки. Поведение каждого животного сначала оценивали в этом контрольном состоянии (см. анализ данных ниже). Чтобы проверить эффективность ряда мотиваторов, в пустую камеру добавили мотиватор, превратив ее в камеру вознаграждения (рис. 1С). Для контрольного теста каждое животное помещали в стартовую камеру и наблюдали в течение одной попытки продолжительностью 2 мин. Регистрировали время, проведенное в каждой камере, и задержку до входа в пустую камеру. Затем это сравнивали с производительностью каждого животного с мотиватором, присутствующим в камере вознаграждения. Всех животных тестировали с одним мотиватором (например, грелкой) в течение одной пробы продолжительностью 2 мин перед переключением на следующий мотиватор, а мотиваторы тестировали в следующем порядке: грелка, затемненная камера (защищенная от внешнего света черной конструкцией).
Те же самые восемь животных, которые использовались при тестировании первой конструкции лабиринта, использовались для определения подходящих мотиваторов для голых землекопов. Сначала в качестве контроля стартовую камеру соединяли с отдельной пустой камерой с помощью Г-образной соединительной трубки. Поведение каждого животного сначала оценивали в этом контрольном состоянии (см. анализ данных ниже). Чтобы проверить эффективность ряда мотиваторов, в пустую камеру добавили мотиватор, превратив ее в камеру вознаграждения (рис. 1С). Для контрольного теста каждое животное помещали в стартовую камеру и наблюдали в течение одной попытки продолжительностью 2 мин. Регистрировали время, проведенное в каждой камере, и задержку до входа в пустую камеру. Затем это сравнивали с производительностью каждого животного с мотиватором, присутствующим в камере вознаграждения. Всех животных тестировали с одним мотиватором (например, грелкой) в течение одной пробы продолжительностью 2 мин перед переключением на следующий мотиватор, а мотиваторы тестировали в следующем порядке: грелка, затемненная камера (защищенная от внешнего света черной конструкцией). бумага), еда (сладкий картофель, банановое детское питание и ProNutro — все предметы, помещенные вместе в пищевую камеру, для всех протестированных животных), подстилка для домашней колонии и подстилка для чужой колонии (таблица 2). Стартовую и пустую камеры или камеры вознаграждения протирали 70% изопропиловым этанолом между сеансами тестирования с разными животными. Полученный в результате оптимизированный мотиватор, представляющий собой комбинацию подстилки для домашней колонии и нагревательной лампы, был использован для камеры непрерывного вознаграждения в лабиринте.
бумага), еда (сладкий картофель, банановое детское питание и ProNutro — все предметы, помещенные вместе в пищевую камеру, для всех протестированных животных), подстилка для домашней колонии и подстилка для чужой колонии (таблица 2). Стартовую и пустую камеры или камеры вознаграждения протирали 70% изопропиловым этанолом между сеансами тестирования с разными животными. Полученный в результате оптимизированный мотиватор, представляющий собой комбинацию подстилки для домашней колонии и нагревательной лампы, был использован для камеры непрерывного вознаграждения в лабиринте.
Таблица 2. Оценка мотиваторов камеры вознаграждения.
Определение ранга доминирования
Ранг в социальной иерархии определялся с использованием парадигмы тестирования доминирования, смоделированной по аналогии с предыдущими исследованиями на голых землекопах (Clarke and Faulkes, 1997, 1998). Все животные, используемые в непрерывной версии лабиринта, были отмечены их идентификационными номерами на спине с использованием перманентного маркера, чтобы можно было визуально отслеживать поведение во время тестирования доминирования. Два исследователя поместили по одному голому землекопу в отдельные круглые камеры. Обе камеры были соединены через прозрачную трубку из поликарбоната. Животные были ориентированы на открытые концы трубки таким образом, чтобы они встречались лицом к лицу, когда они двигались в прямом направлении (рис. 1D). Столкновения оценивались только в том случае, если одно животное проходило над другим в пробирке. Тестовая сессия закончилась после того, как были записаны три случая поведения передачи лицом к лицу. После каждого сеанса камеры и трубки очищали 70% изопропиловым спиртом. Пары животных были созданы случайным образом, при этом ни одна пара не подвергалась более чем одному сеансу тестирования вместе. Каждое животное подвергалось как минимум трем сеансам тестирования, в каждом из которых использовалась новая пара животных.
Два исследователя поместили по одному голому землекопу в отдельные круглые камеры. Обе камеры были соединены через прозрачную трубку из поликарбоната. Животные были ориентированы на открытые концы трубки таким образом, чтобы они встречались лицом к лицу, когда они двигались в прямом направлении (рис. 1D). Столкновения оценивались только в том случае, если одно животное проходило над другим в пробирке. Тестовая сессия закончилась после того, как были записаны три случая поведения передачи лицом к лицу. После каждого сеанса камеры и трубки очищали 70% изопропиловым спиртом. Пары животных были созданы случайным образом, при этом ни одна пара не подвергалась более чем одному сеансу тестирования вместе. Каждое животное подвергалось как минимум трем сеансам тестирования, в каждом из которых использовалась новая пара животных.
Рейтинг доминирования был определен для животных непрерывного лабиринта с использованием ранее описанных методов с голыми землекопами (Clarke and Faulkes, 1997; Gilbert et al. , 2020). Используя расширение пакета для использования с R Studio (R Core Team, 2021 г.) для расчета рейтинга Эло (Neumann and Kulik, 2020 г.), были введены результаты каждого сеанса тестирования за несколько дней, чтобы получить рейтинг каждого отдельного тестируемого животного. Чем выше рейтинг, тем доминантнее животное.
, 2020). Используя расширение пакета для использования с R Studio (R Core Team, 2021 г.) для расчета рейтинга Эло (Neumann and Kulik, 2020 г.), были введены результаты каждого сеанса тестирования за несколько дней, чтобы получить рейтинг каждого отдельного тестируемого животного. Чем выше рейтинг, тем доминантнее животное.
Экспериментальная парадигма
Все испытания пространственного обучения проводились в течение одного и того же 4-часового временного блока в течение каждого дня тестирования (с 8:00 до 12:00). Все испытания проводились в ярко освещенной поведенческой комнате при температуре 18,5–21 °C и относительной влажности не менее 40 %. Животных заносили в комнату в передаточных камерах и давали им акклиматизироваться в наградной камере лабиринта в течение 5 мин. Затем животных извлекали из камеры для вознаграждения и помещали в заданную точку входа в лабиринт лицом вперед (рис. 1А, В). Голому землекопу разрешили исследовать лабиринт, пока он не достиг комнаты вознаграждения. Испытание считалось завершенным, когда прошло 5 минут или когда животное успешно переместилось в камеру вознаграждения, в зависимости от того, что наступит раньше. Если животное не могло добраться до камеры для вознаграждения до истечения 5 минут, экспериментатор осторожно направлял его в камеру после прекращения испытания, и испытание считалось неудачным. При входе в камеру с наградами у входа был установлен блок, чтобы предотвратить повторный вход в лабиринт. Животному давали 5 мин отдохнуть и исследовать камеру, после чего начиналось следующее испытание. Производительность записывалась с помощью видеокамеры (FHD 1080P, YEHOOVJIANGER) и анализировалась в режиме реального времени с использованием программного обеспечения ANY-maze (Stoelting Co., Wood Dale, IL, United States).
Испытание считалось завершенным, когда прошло 5 минут или когда животное успешно переместилось в камеру вознаграждения, в зависимости от того, что наступит раньше. Если животное не могло добраться до камеры для вознаграждения до истечения 5 минут, экспериментатор осторожно направлял его в камеру после прекращения испытания, и испытание считалось неудачным. При входе в камеру с наградами у входа был установлен блок, чтобы предотвратить повторный вход в лабиринт. Животному давали 5 мин отдохнуть и исследовать камеру, после чего начиналось следующее испытание. Производительность записывалась с помощью видеокамеры (FHD 1080P, YEHOOVJIANGER) и анализировалась в режиме реального времени с использованием программного обеспечения ANY-maze (Stoelting Co., Wood Dale, IL, United States).
Животных подвергали пространственному обучению/обучению в течение трех дней подряд с пятью испытаниями в день, всего пятнадцать испытаний (рис. 2). Между каждым испытанием лабиринт очищали 70% изопропиловым спиртом, чтобы уменьшить влияние обонятельных сигналов на производительность в последующих испытаниях. После каждого дня тестирования лабиринт разбирали и промывали водой с мылом без запаха. Сохранение пространственной памяти тестировали через 1 день и 1 неделю после последнего дня обучения для прерывистого лабиринта и через 1 день, 1 неделю и 1 месяц для непрерывного лабиринта (рис. 2). В дни проб памяти животное свободно исследовало лабиринт в течение одной попытки продолжительностью 5 минут или до тех пор, пока не достигало камеры вознаграждения, в зависимости от того, что произошло раньше. В испытаниях с памятью камера вознаграждения содержала те же мотиваторы, что и в испытаниях на обучение. Если животное не могло пройти лабиринт в течение отведенных 5 минут, его возвращали в колонию без направления в камеру вознаграждения.
После каждого дня тестирования лабиринт разбирали и промывали водой с мылом без запаха. Сохранение пространственной памяти тестировали через 1 день и 1 неделю после последнего дня обучения для прерывистого лабиринта и через 1 день, 1 неделю и 1 месяц для непрерывного лабиринта (рис. 2). В дни проб памяти животное свободно исследовало лабиринт в течение одной попытки продолжительностью 5 минут или до тех пор, пока не достигало камеры вознаграждения, в зависимости от того, что произошло раньше. В испытаниях с памятью камера вознаграждения содержала те же мотиваторы, что и в испытаниях на обучение. Если животное не могло пройти лабиринт в течение отведенных 5 минут, его возвращали в колонию без направления в камеру вознаграждения.
Рис. 2. Хронология пространственного обучения и тестирования памяти. Как для прерывистого лабиринта, так и для непрерывного лабиринта животные подвергались 3-дневной поведенческой тренировке, состоящей из пяти проб в день. Через один день, 1 неделю и 1 месяц после последнего дня обучения животные подвергались однократному испытанию памяти. * Испытание пробы памяти в течение одного месяца проводилось для непрерывного лабиринта, но не проводилось для прерывистого лабиринта из-за отсутствия кривой обучения (см. рис. 6).
* Испытание пробы памяти в течение одного месяца проводилось для непрерывного лабиринта, но не проводилось для прерывистого лабиринта из-за отсутствия кривой обучения (см. рис. 6).
Анализ данных
Парная выборка t — тесты были впервые использованы для изучения эффективности различных мотиваторов для использования в камере вознаграждения. P -значения были сначала рассчитаны с использованием необработанных значений для сравнения задержки в камере вознаграждения и задержки в пустой камере (рис. 3А), а также времени пребывания в камере вознаграждения и времени пребывания в пустой камере (рис. 3В). Соотношение времени ожидания для вознаграждения (DTR R ) и условия контроля с пустой камерой (DTR E ) были рассчитаны следующим образом, где DT S представляет собой время пребывания в стартовой камере с вознаграждением (DT SR ) по сравнению с контрольными условиями с пустой камерой (DT SE ), а DT представляет время пребывания в финишной камере между контролем (DT E , время пребывания в пустой камере) и условиями вознаграждения (DT R , время пребывания в награде камера):
DTR=R(DT-SRDT)R/DTSR
DTR=E(DT-SEDT)E/DTSE
Рисунок 3. Эффективность различных мотиваторов для голых землекопов. (A) Из протестированных мотиваторов подстилка для домашней колонии, подстилка для чужой колонии и тепло приводили к самым коротким задержкам входа в камеру вознаграждения по сравнению с пустой камерой. (B) Подстилка как для домашней, так и для чужой колонии привела к наибольшему увеличению времени пребывания в камере вознаграждения по сравнению со стартовой камерой. (C) При оценке коэффициента времени пребывания животные проводили относительно больше времени в камере вознаграждения по сравнению с камерой запуска, когда присутствовала либо домашняя, либо чужая подстилка для колонии. Отрицательный процент указывает на большее время, проведенное в камере вознаграждения, чем в начальной камере. *Указывает на значительную ( p < 0,05) разницу со значением для пустой камеры. с, секунды.
Эффективность различных мотиваторов для голых землекопов. (A) Из протестированных мотиваторов подстилка для домашней колонии, подстилка для чужой колонии и тепло приводили к самым коротким задержкам входа в камеру вознаграждения по сравнению с пустой камерой. (B) Подстилка как для домашней, так и для чужой колонии привела к наибольшему увеличению времени пребывания в камере вознаграждения по сравнению со стартовой камерой. (C) При оценке коэффициента времени пребывания животные проводили относительно больше времени в камере вознаграждения по сравнению с камерой запуска, когда присутствовала либо домашняя, либо чужая подстилка для колонии. Отрицательный процент указывает на большее время, проведенное в камере вознаграждения, чем в начальной камере. *Указывает на значительную ( p < 0,05) разницу со значением для пустой камеры. с, секунды.
Для DTR R по сравнению с DTR 9 также был проведен парный тест t .0085 E (рис. 3C). Бонферрони исправил p значений, которые использовались для определения значимости. Сводные результаты парной выборки t -тестов можно найти в таблице 2.
3C). Бонферрони исправил p значений, которые использовались для определения значимости. Сводные результаты парной выборки t -тестов можно найти в таблице 2.
Для непрерывного лабиринта были проанализированы задержка до камеры вознаграждения и пройденное расстояние для изучения пространственного обучения и производительности памяти (анализ адаптирован из Vallianatou et al. др., 2020). Для животных с отсутствующими точками данных в одном испытании недостающие точки были линейно интерполированы путем усреднения значений для испытания, предшествующего отсутствующей точке, и испытания, следующего за отсутствующей точкой (например, точка данных испытания 4 была интерполирована путем усреднения значений животное в испытании 3 и испытании 5). Чтобы исследовать различия в фазе обучения в непрерывном лабиринте, мы использовали два разных многомерных факторных MANOVA с повторными измерениями. Эти модели включали массу тела и возраст как ковариаты; пол и колония как фиксированные факторы; латентность и расстояние как зависимые меры. Эти модели использовались для определения любых основных воздействий или взаимодействий на производительность аналогично предыдущим исследованиям (Costanzo et al., 2009).; Oosthuizen и др., 2013; Остуйзен, 2020). В первых повторных измерениях MANOVA использовала как «День», так и «Испытание» в качестве внутрисубъектных факторов, при этом каждый день был разделен на соответствующие 5 испытаний. Вторая модель использовалась для простоты интерпретации и моделировала только Испытания без разделения по дням. Во второй модели испытания были проанализированы как испытания с 1 по 15, а не с 1 по 3, каждое с испытанием с 1 по 5. Расчетные предельные средние значения и доверительные интервалы из этих моделей использовались для построения графика скорректированных значений производительности (см. рис. 5, 6). Эти модели с поправкой на ковариаты для массы тела = 54,60 г и возраста = 2,9.4 года. Повторное измерение MANOVA также проводилось отдельно для каждой колонии, чтобы включить рейтинг доминирования в качестве ковариации.
Эти модели использовались для определения любых основных воздействий или взаимодействий на производительность аналогично предыдущим исследованиям (Costanzo et al., 2009).; Oosthuizen и др., 2013; Остуйзен, 2020). В первых повторных измерениях MANOVA использовала как «День», так и «Испытание» в качестве внутрисубъектных факторов, при этом каждый день был разделен на соответствующие 5 испытаний. Вторая модель использовалась для простоты интерпретации и моделировала только Испытания без разделения по дням. Во второй модели испытания были проанализированы как испытания с 1 по 15, а не с 1 по 3, каждое с испытанием с 1 по 5. Расчетные предельные средние значения и доверительные интервалы из этих моделей использовались для построения графика скорректированных значений производительности (см. рис. 5, 6). Эти модели с поправкой на ковариаты для массы тела = 54,60 г и возраста = 2,9.4 года. Повторное измерение MANOVA также проводилось отдельно для каждой колонии, чтобы включить рейтинг доминирования в качестве ковариации. Эти анализы, специфичные для колоний, были проведены, потому что между животными из разных колоний нельзя проводить тестирование доминирования, поскольку это тестирование приводит к смертельным столкновениям. В моделях для конкретных колоний в качестве ковариант использовались масса тела, возраст и ранжирование доминирования; секс как фиксированный фактор; латентность и расстояние как зависимые меры. Эти анализы также были выполнены двумя отдельными способами: один с двумя повторными измерениями «День» и «Испытание», а второй с «Испытаниями» в качестве единственного внутрисубъектного фактора. Колония Расчетные предельные средние значения были рассчитаны с использованием: Масса = 54,63 г, Возраст = 2,35 года, Ранг = 957.40. Расчетные предельные средние значения для колонии B были рассчитаны с использованием: масса = 54,56 г, возраст = 3,49 года, ранг = 1002,96. Все вышеупомянутые статистические анализы также проводились как отдельные повторные измерения ANOVA со скоростью в качестве зависимой переменной для дополнения анализа латентности и расстояния.
Эти анализы, специфичные для колоний, были проведены, потому что между животными из разных колоний нельзя проводить тестирование доминирования, поскольку это тестирование приводит к смертельным столкновениям. В моделях для конкретных колоний в качестве ковариант использовались масса тела, возраст и ранжирование доминирования; секс как фиксированный фактор; латентность и расстояние как зависимые меры. Эти анализы также были выполнены двумя отдельными способами: один с двумя повторными измерениями «День» и «Испытание», а второй с «Испытаниями» в качестве единственного внутрисубъектного фактора. Колония Расчетные предельные средние значения были рассчитаны с использованием: Масса = 54,63 г, Возраст = 2,35 года, Ранг = 957.40. Расчетные предельные средние значения для колонии B были рассчитаны с использованием: масса = 54,56 г, возраст = 3,49 года, ранг = 1002,96. Все вышеупомянутые статистические анализы также проводились как отдельные повторные измерения ANOVA со скоростью в качестве зависимой переменной для дополнения анализа латентности и расстояния. Эти модели проводились отдельно, поскольку скорость равна расстоянию, деленному на задержку. Поправку Бонферрони использовали для апостериорных попарных сравнений для всех эффектов дня и испытания.
Эти модели проводились отдельно, поскольку скорость равна расстоянию, деленному на задержку. Поправку Бонферрони использовали для апостериорных попарных сравнений для всех эффектов дня и испытания.
Проценты производительности в испытаниях памяти были рассчитаны для обеих конфигураций лабиринта (непрерывной и непрерывной), сравнивая производительность (латентность и пройденное расстояние) от отдельных дней проверки памяти до последнего дня обучения. Задержка и пройденное расстояние были проанализированы как для непрерывного, так и для прерывистого дизайна лабиринта, тогда как ошибки были проанализированы только для прерывистого дизайна лабиринта. Они были рассчитаны таким образом, что значение из испытания памяти (1, 2 или 3) было разделено на испытание в последний тренировочный день (испытание 5 в тренировочный день 3; рисунок 2 и таблица 3). MANOVA с повторными измерениями, в котором показатели задержки и расстояния используются в качестве зависимых показателей; масса тела и возраст как ковариаты; пол и колония как фиксированные факторы; и испытание памяти (1, 2 или 3) как внутрисубъектный фактор. Этот анализ проводился для каждой колонии отдельно и в совокупности. Этот набор статистических анализов для пробных испытаний памяти также был проведен как повторные измерения ANOVA со скоростью в качестве зависимой переменной вместо задержки и расстояния.
Этот анализ проводился для каждой колонии отдельно и в совокупности. Этот набор статистических анализов для пробных испытаний памяти также был проведен как повторные измерения ANOVA со скоростью в качестве зависимой переменной вместо задержки и расстояния.
Таблица 3. Сравнение производительности между обеими конфигурациями лабиринта для оценки пространственной памяти.
Результаты
Проверка относительной эффективности ряда поведенческих мотиваторов у голых землекопов
Мы протестировали различные мотиваторы, чтобы оптимизировать последующую производительность тестирования в пространственном лабиринте (рис. 1C). При использовании парных тестов t для исходных значений латентности, времени пребывания и соотношения времени пребывания по сравнению с пустой камерой аналогичные мотиваторы оказались значительными (таблица 2 и рисунок 3). Латентный период усиливался при наличии подстилки домашней колонии [ t (7) = 5,904, p = 0,003] или залегание инородных колоний [ t (7) = 5,352, p = 0,005], с приближением тепла к значимости [ t (7,) = 3,21 р = 0,075]. Отношение времени пребывания не было значимым для любого тестируемого мотиватора, но время пребывания было значительно больше, когда подстилка домашней семьи [ t (7) = -9,623, p <0,001] или подстилка иностранной семьи [ t (7) = −7,904, p < 0,001] присутствовали в наградной камере. Основываясь на этих наборах анализов, тепло и наличие любого типа постельных принадлежностей (иностранные или домашние) были наиболее эффективными мотиваторами при независимом представлении. Таким образом, и тепло, и домашняя подстилка были выбраны для камеры вознаграждения в последующем непрерывном тестировании лабиринта. Подстилка для чужой колонии, хотя и также эффективная, не была выбрана из-за потенциального риска нападения на экспериментальное животное при повторном введении в домашнюю колонию из-за запаха злоумышленника (Braude et al., 2021).
Отношение времени пребывания не было значимым для любого тестируемого мотиватора, но время пребывания было значительно больше, когда подстилка домашней семьи [ t (7) = -9,623, p <0,001] или подстилка иностранной семьи [ t (7) = −7,904, p < 0,001] присутствовали в наградной камере. Основываясь на этих наборах анализов, тепло и наличие любого типа постельных принадлежностей (иностранные или домашние) были наиболее эффективными мотиваторами при независимом представлении. Таким образом, и тепло, и домашняя подстилка были выбраны для камеры вознаграждения в последующем непрерывном тестировании лабиринта. Подстилка для чужой колонии, хотя и также эффективная, не была выбрана из-за потенциального риска нападения на экспериментальное животное при повторном введении в домашнюю колонию из-за запаха злоумышленника (Braude et al., 2021).
Пространственное обучение у голых землекопов
Для предварительного набора данных, собранных с использованием дизайна прерывистого лабиринта ( n = 8; рис. 1A), кривые обучения не были очевидны в ходе обучающих испытаний для задержки в камере вознаграждения ( Рисунок 4A), общее количество сделанных ошибок (неправильных поворотов) (Рисунок 4B) или общее пройденное расстояние (Рисунок 4C). Частота отказов также была высокой (таблица 1). Плохая работа голых землекопов в этой прерывистой конструкции лабиринта, наряду с наблюдением, что прерывистая конструкция лабиринта могла создать путаницу (т. землекопы для демонстрации пространственного обучения и улучшения памяти), привели нас к перепроектированию лабиринта в виде непрерывной схемы без тупиков.
1A), кривые обучения не были очевидны в ходе обучающих испытаний для задержки в камере вознаграждения ( Рисунок 4A), общее количество сделанных ошибок (неправильных поворотов) (Рисунок 4B) или общее пройденное расстояние (Рисунок 4C). Частота отказов также была высокой (таблица 1). Плохая работа голых землекопов в этой прерывистой конструкции лабиринта, наряду с наблюдением, что прерывистая конструкция лабиринта могла создать путаницу (т. землекопы для демонстрации пространственного обучения и улучшения памяти), привели нас к перепроектированию лабиринта в виде непрерывной схемы без тупиков.
Рис. 4. Непрерывное обучение в пространственном лабиринте у голых землекопов. Предварительные данные были собраны ( n = 8) с использованием прерывистой модифицированной версии лабиринта Хебба-Вильямса (см. рис. 1А) для исследования возможностей пространственного обучения и памяти у голых землекопов в ходе 15 тренировочных испытаний (5 испытаний в день). на 3 дня). (A) Задержка достижения камеры вознаграждения усреднялась по животным для каждого испытания обучения/обучения. Непрерывное выполнение лабиринта показало очень постепенную кривую обучения задержки с тенденцией к уменьшению задержки в испытаниях, но большую вариабельность в индивидуальной производительности. (B) Среднее количество входов в зону ошибки было нанесено на график для всех животных, протестированных в ходе испытаний по обучению/обучению. Для прерывистого лабиринта не было кривой обучения уменьшению ошибок. (C) Общее расстояние, пройденное до достижения камеры вознаграждения, усреднялось по животным для каждого испытания и наносилось на график для всех испытаний обучения/обучения. Непрерывное прохождение лабиринта показало большую вариабельность индивидуальной производительности и отсутствие кривой обучения, связанной с уменьшением пройденного расстояния (оптимизация пути к камере вознаграждения). м, м; с, секунды.
(A) Задержка достижения камеры вознаграждения усреднялась по животным для каждого испытания обучения/обучения. Непрерывное выполнение лабиринта показало очень постепенную кривую обучения задержки с тенденцией к уменьшению задержки в испытаниях, но большую вариабельность в индивидуальной производительности. (B) Среднее количество входов в зону ошибки было нанесено на график для всех животных, протестированных в ходе испытаний по обучению/обучению. Для прерывистого лабиринта не было кривой обучения уменьшению ошибок. (C) Общее расстояние, пройденное до достижения камеры вознаграждения, усреднялось по животным для каждого испытания и наносилось на график для всех испытаний обучения/обучения. Непрерывное прохождение лабиринта показало большую вариабельность индивидуальной производительности и отсутствие кривой обучения, связанной с уменьшением пройденного расстояния (оптимизация пути к камере вознаграждения). м, м; с, секунды.
Рис. 5. Эффективность обучения в непрерывном пространственном лабиринте у голых землекопов. Данные были собраны ( n = 27) с использованием конструкции непрерывного лабиринта (см. рис. 1B) для исследования пространственного обучения и возможностей памяти у голых землекопов в ходе 15 тренировочных испытаний (5 испытаний в день в течение 3 дней). (A) Средняя задержка в камере вознаграждения выявила четкую кривую обучения в ходе испытаний по обучению/обучению. Сравнение испытания 1 с испытаниями 2–15 выявило значительное снижение латентности в испытании 3. (B) Среднее общее пройденное расстояние также продемонстрировало кривую обучения в процессе обучения/учебных испытаний. Сравнение испытания 1 с испытаниями 2–15 выявило значительное уменьшение расстояния в испытании 3. (C) Средняя пройденная скорость значительно увеличилась во время тренировочных испытаний в испытании 6. * p < 0,05 по сравнению с испытанием 1. м, метров; с, секунды.
Данные были собраны ( n = 27) с использованием конструкции непрерывного лабиринта (см. рис. 1B) для исследования пространственного обучения и возможностей памяти у голых землекопов в ходе 15 тренировочных испытаний (5 испытаний в день в течение 3 дней). (A) Средняя задержка в камере вознаграждения выявила четкую кривую обучения в ходе испытаний по обучению/обучению. Сравнение испытания 1 с испытаниями 2–15 выявило значительное снижение латентности в испытании 3. (B) Среднее общее пройденное расстояние также продемонстрировало кривую обучения в процессе обучения/учебных испытаний. Сравнение испытания 1 с испытаниями 2–15 выявило значительное уменьшение расстояния в испытании 3. (C) Средняя пройденная скорость значительно увеличилась во время тренировочных испытаний в испытании 6. * p < 0,05 по сравнению с испытанием 1. м, метров; с, секунды.
Рис. 6. Непрерывные кривые обучения пространственного лабиринта с использованием расчетных предельных средних значений. (А) Задержка в камере вознаграждения была значительно уменьшена в среднем в испытаниях 2, 3, 4 и 5 по сравнению с испытанием 1 в дни 1–3, демонстрируя основной эффект испытания независимо от дня задержки. (B) Средняя задержка для каждого дня обучения показала, что животные добирались до камеры вознаграждения за значительно меньшее время в дни 2 и 3 по сравнению с днем 1. (C) Животные продемонстрировали значительный эффект обучения при сравнении расстояния, пройденного в испытаниях 2. , 3 и 5 по сравнению с испытанием 1 в дни 1–3, демонстрируя основной эффект испытания, не зависящий от дня, для расстояния. (D) Животные прошли значительно меньшее расстояние в день 2 по сравнению с днем 1. Коварианты, появляющиеся в модели, оценивались по следующим значениям: масса = 54,60, возраст = 2,94.* p < 0,05. м, м; с, секунды.
(А) Задержка в камере вознаграждения была значительно уменьшена в среднем в испытаниях 2, 3, 4 и 5 по сравнению с испытанием 1 в дни 1–3, демонстрируя основной эффект испытания независимо от дня задержки. (B) Средняя задержка для каждого дня обучения показала, что животные добирались до камеры вознаграждения за значительно меньшее время в дни 2 и 3 по сравнению с днем 1. (C) Животные продемонстрировали значительный эффект обучения при сравнении расстояния, пройденного в испытаниях 2. , 3 и 5 по сравнению с испытанием 1 в дни 1–3, демонстрируя основной эффект испытания, не зависящий от дня, для расстояния. (D) Животные прошли значительно меньшее расстояние в день 2 по сравнению с днем 1. Коварианты, появляющиеся в модели, оценивались по следующим значениям: масса = 54,60, возраст = 2,94.* p < 0,05. м, м; с, секунды.
Обе колонии (колония A и колония B) были проанализированы вместе на максимальную мощность с дизайном непрерывного лабиринта ( n = 27; рис. 1B) и с использованием повторных измерений MANOVA, которые не моделировали эффект тестирования в отдельные дни. для простоты интерпретации. Этот анализ выявил четкую кривую обучения в 15 тренировочных испытаниях (рис. 5А), которая отсутствовала в прерывистом лабиринте. Отмечалось многофакторное влияние количества проб на производительность [9].0023 F (28, 588) = 3,074, p < 0,001, рисунок 5А и дополнительная таблица 1]. Многофакторный эффект Trial был обусловлен основными эффектами как задержки, так и расстояния. Повторные попарные сравнения испытаний 1 с испытаниями 2–15 показали, что задержка ( с поправкой Бонферрони p <0,05, рисунок 5A и дополнительная таблица 1) и расстояние ( с поправкой Бонферрони p <0,05, рисунок 5B и дополнительная таблица 1) значительно уменьшились. к 3-й попытке, тогда как скорость значительно увеличилась к 6-й попытке (Бонферрони исправил p <0,05, рисунок 5C и дополнительная таблица 3). Кроме того, у самок животных было значительно большее увеличение скорости обучения, чем у самцов [ F (14, 294) = 2,811, p = 0,001, дополнительная таблица 3], хотя существенных различий между полами не было.
1B) и с использованием повторных измерений MANOVA, которые не моделировали эффект тестирования в отдельные дни. для простоты интерпретации. Этот анализ выявил четкую кривую обучения в 15 тренировочных испытаниях (рис. 5А), которая отсутствовала в прерывистом лабиринте. Отмечалось многофакторное влияние количества проб на производительность [9].0023 F (28, 588) = 3,074, p < 0,001, рисунок 5А и дополнительная таблица 1]. Многофакторный эффект Trial был обусловлен основными эффектами как задержки, так и расстояния. Повторные попарные сравнения испытаний 1 с испытаниями 2–15 показали, что задержка ( с поправкой Бонферрони p <0,05, рисунок 5A и дополнительная таблица 1) и расстояние ( с поправкой Бонферрони p <0,05, рисунок 5B и дополнительная таблица 1) значительно уменьшились. к 3-й попытке, тогда как скорость значительно увеличилась к 6-й попытке (Бонферрони исправил p <0,05, рисунок 5C и дополнительная таблица 3). Кроме того, у самок животных было значительно большее увеличение скорости обучения, чем у самцов [ F (14, 294) = 2,811, p = 0,001, дополнительная таблица 3], хотя существенных различий между полами не было. для задержки и расстояния между испытаниями.
для задержки и расстояния между испытаниями.
При использовании модели, которая включала в себя отдельные дни тестирования, были получены аналогичные результаты. Наблюдался многомерный эффект Trial [ F (8, 168) = 3,732, p < 0,001, дополнительная таблица 2], обусловленный значительными основными эффектами испытания на обе задержки [ F (4, 84) = 5,494, , , , , = 0,001, дополнительная таблица 2] и расстояние [ F (4, 84) = 2,809, p = 0,031, дополнительная таблица 2], но не скорость [ F (4, 84)6 = , p = 0,051, дополнительная таблица 4]. Парные сравнения с поправкой Бонферрони показали, что животные путешествовали значительно меньше во время испытаний 2, 3 и 5 в любой день по сравнению с испытанием 1 (-1,888, -2,4843, -1,9).10 и −2,574 м соответственно; р < 0,05; Рисунок 6C и дополнительная таблица 2). Точно так же задержка значительно уменьшилась от испытания 1 до испытаний 2, 3 и 5 (соответствующие изменения -43,515, -61,82 и -72,84 с; p <0,05; рисунок 6A и дополнительная таблица 2).

Не было многомерного эффекта Day, независимо от номера испытания. Также не было выявлено одномерного основного влияния Дня на задержку, скорость или расстояние, пройденное для достижения камеры вознаграждения. Однако попарные сравнения показали, что расстояние значительно уменьшилось между 1 и 2 днями тренировки (-1,768 м, Бонферрони исправил 9).0023 р = 0,002; Рисунок 6D и дополнительная таблица 2). Не было значительного уменьшения расстояния между пройденным расстоянием в 1-й и 3-й дни тренировки. Средняя задержка между испытаниями в 1-й день периода обучения была значительно больше, чем латентность во 2-й день (70,376 с, p <0,05; рисунок 6B и дополнительная таблица 2) и в 3-й день (76,556 с, p <0,05); Рисунок 6B и дополнительная таблица 2).
Было значимое многомерное взаимодействие Day by Trial [ F (16, 36) = 3,534, p < 0,001, дополнительная таблица 2], обусловленные значительным взаимодействием для обеих задержек [ F (8, 168) = 6,006, p <дополнительной таблицы
2] и расстояние [ F (8, 168) = 4,084, p <0,001, дополнительная таблица 2]. Также наблюдалось значительное взаимодействие Day и Trial по скорости [ F (8, 168) = 2,40, p = 0,018, дополнительная таблица 4]. Как задержка, так и расстояние продемонстрировали наибольшие улучшения производительности в испытаниях в первый день обучения, при этом улучшения производительности стабилизировались в испытаниях в дни 2 и 3, показывая, что большая часть обучения происходила в день 1.
Также наблюдалось значительное взаимодействие Day и Trial по скорости [ F (8, 168) = 2,40, p = 0,018, дополнительная таблица 4]. Как задержка, так и расстояние продемонстрировали наибольшие улучшения производительности в испытаниях в первый день обучения, при этом улучшения производительности стабилизировались в испытаниях в дни 2 и 3, показывая, что большая часть обучения происходила в день 1.
Не было многомерного взаимодействия между днем и массой тела, однако было одномерное влияние на латентность [ F (2, 42) = 4,137, p = 0,023, дополнительная таблица 2], но не на расстояние . Более крупные животные имели более высокие задержки, чтобы добраться до камеры вознаграждения в первые дни обучения, и эти различия уменьшались в течение дня, несмотря на то, что все животные преодолевали сопоставимые расстояния.
Отсутствовало многовариантное взаимодействие Trial by Age. Однако было выявлено одномерное влияние Испытания по возрасту на расстояние [ F (4, 84) = 2,580, p = 0,043, дополнительная таблица 2], что указывает на лучшую производительность для более молодых животных в испытаниях 2 и 3, но для более старых животных в испытаниях 4 и 5. многомерный трехсторонний день по испытанию по взаимодействию с массой тела [ F (16, 336) = 1,988, p = 0,013, дополнительная таблица 2], обусловленный значительными взаимодействиями по обеим задержкам [ F (8, 168) = 3,245, p = 0,002, дополнительная таблица 2] и расстояние [ F (8, 168) = 2,398, p = 0,018, дополнительная таблица 2]. Этот эффект указывал на то, что на форму кривой обучения по дням и испытаниям влияла масса тела: более мелкие животные изначально демонстрировали более быстрое обучение, с более изменчивыми результатами в более поздних тренировочных испытаниях.
многомерный трехсторонний день по испытанию по взаимодействию с массой тела [ F (16, 336) = 1,988, p = 0,013, дополнительная таблица 2], обусловленный значительными взаимодействиями по обеим задержкам [ F (8, 168) = 3,245, p = 0,002, дополнительная таблица 2] и расстояние [ F (8, 168) = 2,398, p = 0,018, дополнительная таблица 2]. Этот эффект указывал на то, что на форму кривой обучения по дням и испытаниям влияла масса тела: более мелкие животные изначально демонстрировали более быстрое обучение, с более изменчивыми результатами в более поздних тренировочных испытаниях.
Не было ни многомерного воздействия Day by Trial by Sex на производительность, ни одномерного воздействия на задержку и расстояние. Однако имело место значительное трехстороннее одномерное взаимодействие по скорости. Самки животных увеличивали скорость во время обучающих испытаний значительно больше, чем самцы [9].0023 F (8, 168) = 3,443, p = 0,001, дополнительная таблица 4].
Никакие другие многомерные или одномерные эффекты или взаимодействия (день по возрасту, день по полу или день по колонии, испытание по массе, испытание по полу и испытание по колонии) не были значительными для задержки достижения наградной камеры, пройденного расстояния, или скорость.
Чтобы включить рейтинг доминирования (специфический показатель колонии) в GLM, набор данных был разделен по колониям для анализа. Не было значительного влияния ранга доминирования на задержку или расстояние между испытаниями и днями, а также не было никаких взаимодействий ранга доминирования с какими-либо другими переменными в модели (подробную статистику см. в дополнительных таблицах 1, 2). Скорость передвижения варьировалась в зависимости от ранга доминирования для колонии B, при этом подчиненные животные перемещались быстрее в испытаниях, чем доминирующие животные (подробную статистику см. в дополнительных таблицах 3, 4).
Эти результаты показывают, что голые землекопы демонстрировали значительное пространственное обучение в непрерывном лабиринте. Основная часть улучшений производительности произошла в течение первых 3 испытаний, что указывает на быстрое пространственное обучение. Расчеты как задержки, так и расстояния были информативны при оценке результатов пространственного обучения в этой конкретной конфигурации лабиринта у голых землекопов, что позволяет предположить, что голые землекопы смогли быстро научиться ориентироваться в непрерывном дизайне лабиринта.
Основная часть улучшений производительности произошла в течение первых 3 испытаний, что указывает на быстрое пространственное обучение. Расчеты как задержки, так и расстояния были информативны при оценке результатов пространственного обучения в этой конкретной конфигурации лабиринта у голых землекопов, что позволяет предположить, что голые землекопы смогли быстро научиться ориентироваться в непрерывном дизайне лабиринта.
Пространственная память у голых землекопов
Средняя производительность была проанализирована в тестах памяти как для прерывистого лабиринта, так и для непрерывного лабиринта (таблица 3). Не было никаких существенных различий между последним испытанием в последний учебный день и испытаниями памяти 1, 2 или 3 в оценке латентного периода (рис. 7А) или пройденного расстояния (рис. 7В), а также каких-либо влияний возраста, пола, ранжирования, или масса. Это свидетельствует о том, что пространственная память сохранялась у голых землекопов не менее 1 мес.
Рисунок 7. Производительность памяти в непрерывном пространственном лабиринте у голых землекопов. Животных тестировали на сохранение пространственной памяти через 1 день, 1 неделю и 1 месяц (проба памяти 1, 2 и 3 соответственно) после обучения. Пунктирные линии показывают точку эквивалентности результатов в последний тренировочный день. (A) Задержка между испытаниями проб памяти, при этом показатели задержки пробы памяти показаны в процентах от последней попытки в последний день обучения на этапе обучения тестирования. Никаких существенных различий не было обнаружено, что указывает на сохранение пространственной памяти, хотя это могло быть искажено присутствующей высокой изменчивостью. (B) Общее расстояние, пройденное в ходе испытаний проб памяти, при этом расстояние, пройденное во время указанного испытания пробы памяти, показано в процентах от расстояния, пройденного во время последней попытки последнего тренировочного дня. Никаких существенных ухудшений производительности не было отмечено в течение трех временных точек.
Производительность памяти в непрерывном пространственном лабиринте у голых землекопов. Животных тестировали на сохранение пространственной памяти через 1 день, 1 неделю и 1 месяц (проба памяти 1, 2 и 3 соответственно) после обучения. Пунктирные линии показывают точку эквивалентности результатов в последний тренировочный день. (A) Задержка между испытаниями проб памяти, при этом показатели задержки пробы памяти показаны в процентах от последней попытки в последний день обучения на этапе обучения тестирования. Никаких существенных различий не было обнаружено, что указывает на сохранение пространственной памяти, хотя это могло быть искажено присутствующей высокой изменчивостью. (B) Общее расстояние, пройденное в ходе испытаний проб памяти, при этом расстояние, пройденное во время указанного испытания пробы памяти, показано в процентах от расстояния, пройденного во время последней попытки последнего тренировочного дня. Никаких существенных ухудшений производительности не было отмечено в течение трех временных точек.
Обсуждение
В ходе настоящего исследования был создан индивидуальный и эффективный дизайн лабиринта для тестирования пространственного обучения и памяти у голых землекопов. Мы предсказали, что животные будут демонстрировать пространственное обучение, достигая камеры вознаграждения быстрее (уменьшается задержка) и более искусно (более короткое общее пройденное расстояние и / или меньше совершенных ошибок, что указывает на оптимизированную навигацию по лабиринту) во время тренировочных испытаний. Это приведет к крутым кривым обучения и резким плато при достижении пиковой производительности. Чтобы эффективно создать лабиринт для оценки этих показателей обучения и памяти, мы внесли целевые изменения в конструкцию лабиринта Хебба-Уильямса для использования у голых землекопов на основе их уникальных сенсорных систем.
Голые землекопы функционально слепы, с низкой остротой зрения, но с некоторыми остатками светочувствительности. Оливарное претектальное ядро, область, участвующая в обнаружении света, мала, но сохраняет свою типичную морфологию (Crish et al. , 2006). В сочетании с сохраненной реакцией сужения зрачков на свет, в дополнение к поведенческим реакциям на обнаружение света (например, замирание при внезапном изменении яркости), голые землекопы сохраняют способность обнаруживать изменения интенсивности света (Hetling et al., 2005). но в остальном слабовидящие. Считается, что такое снижение зрительных возможностей сохраняет сенсорные ресурсы в пользу других модальностей (например, тактильных), которые более полезны в подземных туннельных системах, в которых отсутствует множество значимых визуальных сигналов, которые облегчили бы такие действия, как навигация. Поскольку традиционные протоколы для пространственных лабиринтов используют визуальные подсказки, были необходимы корректировки, чтобы приспособиться к дефициту зрения у голых землекопов.
, 2006). В сочетании с сохраненной реакцией сужения зрачков на свет, в дополнение к поведенческим реакциям на обнаружение света (например, замирание при внезапном изменении яркости), голые землекопы сохраняют способность обнаруживать изменения интенсивности света (Hetling et al., 2005). но в остальном слабовидящие. Считается, что такое снижение зрительных возможностей сохраняет сенсорные ресурсы в пользу других модальностей (например, тактильных), которые более полезны в подземных туннельных системах, в которых отсутствует множество значимых визуальных сигналов, которые облегчили бы такие действия, как навигация. Поскольку традиционные протоколы для пространственных лабиринтов используют визуальные подсказки, были необходимы корректировки, чтобы приспособиться к дефициту зрения у голых землекопов.
Подземная среда обитания голого землекопа, как полагают, также привела к снижению слуховых способностей. Голые землекопы наиболее чувствительны к частотам от 65 до 12,8 кГц (Heffner, Heffner, 1993; Okanoya et al. , 2018). Голые землекопы демонстрируют плохую локализацию звука в дополнение к меньшим слуховым ядрам ствола мозга, связанным с бинауральной обработкой высокочастотного звука (Heffner and Heffner, 1993). Их улитки характеризуются менее обширной спиральностью (Mason et al., 2016). Эти особенности, наряду с дифференциальными вариациями плотности волосковых клеток в улитке (Barone et al., 2019), все возможные вклады в продемонстрированные у этого вида плохие способности к локализации звука (Heffner and Heffner, 1993). Из-за относительно ограниченных слуховых возможностей голых землекопов слуховые стимулы не были включены в конструкцию пространственного лабиринта, чтобы помочь животным добраться до камеры вознаграждения.
, 2018). Голые землекопы демонстрируют плохую локализацию звука в дополнение к меньшим слуховым ядрам ствола мозга, связанным с бинауральной обработкой высокочастотного звука (Heffner and Heffner, 1993). Их улитки характеризуются менее обширной спиральностью (Mason et al., 2016). Эти особенности, наряду с дифференциальными вариациями плотности волосковых клеток в улитке (Barone et al., 2019), все возможные вклады в продемонстрированные у этого вида плохие способности к локализации звука (Heffner and Heffner, 1993). Из-за относительно ограниченных слуховых возможностей голых землекопов слуховые стимулы не были включены в конструкцию пространственного лабиринта, чтобы помочь животным добраться до камеры вознаграждения.
В отличие от слуховой и зрительной систем соматосенсорная система голых землекопов хорошо развита и включает в себя массив осязательных волосков, распределенных по всему телу и лицу (Crish et al., 2003; Henry et al., 2006). ). Имея около 40 тактильных волосков на каждой стороне тела, голые землекопы демонстрируют сильные ориентировочные реакции на стимуляцию отдельных волосков (Crish et al. , 2003). Эти сенсорные волоски также присутствуют у некоторых представителей Bathyergidae (Bennett and Jarvis, 2004; Bennett et al., 2009).; Walcher et al., 2018), предоставляя сенсорную информацию, компенсирующую нарушения зрения у подземных грызунов (Jarvis and Bennett, 2017). Основываясь на этих сенсорных специализациях, мы построили пространственный лабиринт с трубками, которые были достаточно узкими, чтобы стимулировать тактильные волоски и давать пространственные сигналы. Предыдущие исследования на слепых землекопах продемонстрировали повышение эффективности обучения и памяти при использовании столь же узких проходов, которые давали тактильные сигналы в пространственных лабиринтах (Kimchi and Terkel, 2004).
, 2003). Эти сенсорные волоски также присутствуют у некоторых представителей Bathyergidae (Bennett and Jarvis, 2004; Bennett et al., 2009).; Walcher et al., 2018), предоставляя сенсорную информацию, компенсирующую нарушения зрения у подземных грызунов (Jarvis and Bennett, 2017). Основываясь на этих сенсорных специализациях, мы построили пространственный лабиринт с трубками, которые были достаточно узкими, чтобы стимулировать тактильные волоски и давать пространственные сигналы. Предыдущие исследования на слепых землекопах продемонстрировали повышение эффективности обучения и памяти при использовании столь же узких проходов, которые давали тактильные сигналы в пространственных лабиринтах (Kimchi and Terkel, 2004).
На этапе предварительного сбора данных в этом исследовании мы первоначально реализовали модифицированный дизайн лабиринта Хебба-Уильямса, который был прерывистым (рис. 1А). Однако эта конструкция лабиринта оказалась неэффективной в выявлении истинных способностей к пространственному обучению и памяти у голых землекопов, и при тестировании животных в прерывистом лабиринте не было выявлено никаких кривых обучения (рис. 6). Это может быть связано с отвлечением внимания от тупиков туннеля. Голые землекопы предпочитали копать или прокусывать тупики туннеля, а не искать камеру с вознаграждением, что сильно влияло на показатели пространственного обучения и памяти (например, латентность) в дополнение к влиянию на завершение процесса. лабиринт в течение отведенного времени (т. е. частота неудач, таблица 1). Эти поведенческие наблюдения подтверждаются предыдущими исследованиями, показавшими, что голые землекопы очень заинтересованы в удалении любых блокировок в своих туннельных системах (Brett, 19).91).
6). Это может быть связано с отвлечением внимания от тупиков туннеля. Голые землекопы предпочитали копать или прокусывать тупики туннеля, а не искать камеру с вознаграждением, что сильно влияло на показатели пространственного обучения и памяти (например, латентность) в дополнение к влиянию на завершение процесса. лабиринт в течение отведенного времени (т. е. частота неудач, таблица 1). Эти поведенческие наблюдения подтверждаются предыдущими исследованиями, показавшими, что голые землекопы очень заинтересованы в удалении любых блокировок в своих туннельных системах (Brett, 19).91).
Основываясь на результатах прерывистого лабиринта, мы исследовали мотиваторы, которые хорошо работали бы в паре с совпадающими желаниями кусать и копать в условиях туннеля. Как правило, пространственные лабиринты полагаются на пищевые вознаграждения, чтобы мотивировать животных пройти лабиринт (Олтон и Самуэльсон, 1976; Браун и Кук, 1986; дю Туа и др., 2012). Однако структура колонии голых землекопов затрудняет лишение животных пищи без постоянной изоляции от их колонии, а это, в свою очередь, может отрицательно сказаться на поведенческих характеристиках. Таким образом, необходимо было разработать альтернативные методы мотивации. Голые землекопы были классифицированы как пойкилотермы — плохие регуляторы собственной температуры тела (Яхав и Баффенштейн, 19).92), но это зависит от их состояния активности, и поэтому их можно более точно охарактеризовать как гетеротермические (Kovalzon et al., 2020, 2021). Поэтому в наградную камеру в качестве мотиватора была добавлена тепловая лампа (таблица 2 и рисунок 3). Как роющие грызуны, животные быстро проникали в камеру с вознаграждением, когда подстилка присутствовала, и сразу же начинали копаться в ней и пинать ее по камере. Поскольку домашние и чужие подстилки для колонии оказались эффективными мотиваторами (таблица 2 и рисунок 3), мы включили комбинацию подстилки для домашней колонии и нагревательной лампы в наградной камере для последующего тестирования в лабиринте, подтверждая предыдущие исследования, показывающие желательность домашней подстилки. (О’Риэн и Джарвис, 19 лет.97; Диакон, 2012; Тор и др.
Таким образом, необходимо было разработать альтернативные методы мотивации. Голые землекопы были классифицированы как пойкилотермы — плохие регуляторы собственной температуры тела (Яхав и Баффенштейн, 19).92), но это зависит от их состояния активности, и поэтому их можно более точно охарактеризовать как гетеротермические (Kovalzon et al., 2020, 2021). Поэтому в наградную камеру в качестве мотиватора была добавлена тепловая лампа (таблица 2 и рисунок 3). Как роющие грызуны, животные быстро проникали в камеру с вознаграждением, когда подстилка присутствовала, и сразу же начинали копаться в ней и пинать ее по камере. Поскольку домашние и чужие подстилки для колонии оказались эффективными мотиваторами (таблица 2 и рисунок 3), мы включили комбинацию подстилки для домашней колонии и нагревательной лампы в наградной камере для последующего тестирования в лабиринте, подтверждая предыдущие исследования, показывающие желательность домашней подстилки. (О’Риэн и Джарвис, 19 лет.97; Диакон, 2012; Тор и др. , 2015). Подстилка для чужеродных колоний, хотя и эффективная для увеличения времени, проведенного в камере вознаграждения, и уменьшения задержки в камере вознаграждения, была признана неоптимальной и не использовалась в последующем тестировании лабиринта. Это решение было основано на территориальном характере голых землекопов (O’Riain and Jarvis, 1997; Braude et al., 2021) и риске нападения на экспериментальное животное при реинтродукции в домашнюю колонию из-за запаха незваного гостя. .
, 2015). Подстилка для чужеродных колоний, хотя и эффективная для увеличения времени, проведенного в камере вознаграждения, и уменьшения задержки в камере вознаграждения, была признана неоптимальной и не использовалась в последующем тестировании лабиринта. Это решение было основано на территориальном характере голых землекопов (O’Riain and Jarvis, 1997; Braude et al., 2021) и риске нападения на экспериментальное животное при реинтродукции в домашнюю колонию из-за запаха незваного гостя. .
В нашей переработанной непрерывной версии лабиринта Хебба-Уильямса не было тупиков, которые голые землекопы могли бы интерпретировать как закупорку их туннельных систем (рис. 1В), что привело к снижению частоты отказов (Таблица 1). В отличие от прерывистого лабиринта (рис. 4А), кривые обучения были очевидны в непрерывном лабиринте (рис. 5А). При использовании MANOVA с повторными измерениями как задержка, так и расстояние в испытаниях обучения показали значительные кривые обучения в зависимости от испытаний и дней (рис. 5A, B). Таким образом, текущее исследование продемонстрировало пространственное обучение у голых землекопов с использованием индивидуальной конструкции непрерывного лабиринта с задержкой до камеры вознаграждения и пройденным расстоянием в качестве показателей обучения. Более мелкие животные продемонстрировали более быстрое обучение (дополнительные таблицы 1, 2). Не было многомерного эффекта «День за испытанием по возрасту». Общее влияние возраста на производительность может быть уменьшено из-за сравнительно небольшого возрастного диапазона, проверенного в текущем исследовании (примерно 7 лет), по сравнению с более чем 30-летней продолжительностью жизни содержащихся в неволе голых землекопов (Buffenstein and Craft, 2021; Buffenstein et al. др., 2022). В Day by Trial by Sex взаимодействия ни латентность, ни расстояние между полами существенно не различались. Одно из возможных объяснений отсутствия влияния пола на пространственное обучение и память состоит в том, что эти различия будут присутствовать у видов с половыми различиями в домашних ареалах (Costanzo et al.
5A, B). Таким образом, текущее исследование продемонстрировало пространственное обучение у голых землекопов с использованием индивидуальной конструкции непрерывного лабиринта с задержкой до камеры вознаграждения и пройденным расстоянием в качестве показателей обучения. Более мелкие животные продемонстрировали более быстрое обучение (дополнительные таблицы 1, 2). Не было многомерного эффекта «День за испытанием по возрасту». Общее влияние возраста на производительность может быть уменьшено из-за сравнительно небольшого возрастного диапазона, проверенного в текущем исследовании (примерно 7 лет), по сравнению с более чем 30-летней продолжительностью жизни содержащихся в неволе голых землекопов (Buffenstein and Craft, 2021; Buffenstein et al. др., 2022). В Day by Trial by Sex взаимодействия ни латентность, ни расстояние между полами существенно не различались. Одно из возможных объяснений отсутствия влияния пола на пространственное обучение и память состоит в том, что эти различия будут присутствовать у видов с половыми различиями в домашних ареалах (Costanzo et al. , 2009).), чего у наших голых землекопов нет из-за общей системы нор для самцов и самок. Не было значительного влияния рейтинга доминирования (дополнительные таблицы 1, 2), несмотря на предыдущие публикации, показывающие влияние социальной иерархии на пространственную память (Costanzo et al., 2009).
, 2009).), чего у наших голых землекопов нет из-за общей системы нор для самцов и самок. Не было значительного влияния рейтинга доминирования (дополнительные таблицы 1, 2), несмотря на предыдущие публикации, показывающие влияние социальной иерархии на пространственную память (Costanzo et al., 2009).
При использовании повторных измерений MANOVA-анализа для анализа сохранения в памяти прироста пространственного обучения значительное снижение производительности памяти не было очевидным. Это указывает на хорошее сохранение памяти в течение по крайней мере 1 месяца (таблица 3 и рисунок 7), однако важно отметить, что мотиваторы присутствовали в камере вознаграждения во время испытаний проб памяти, и животные могли следовать сенсорным сигналам, а не явно. используя пространственную память. Эти результаты подтверждаются предыдущими исследованиями, показывающими, что другие виды землекопов не испытывали снижения производительности/удержания пространственной памяти в течение 1 месяца после обучения, превосходя более дальних родственников внутри Rodentia, крысу и степную полевку (Кимчи и Теркель, 2001).
Заключение и дальнейшие направления
Результаты этого исследования позволяют по-новому взглянуть на соображения, которые необходимо учитывать при разработке поведенческих тестов для атипичных моделей животных. При проведении поведенческого тестирования представителей Bathyergidae важно создавать задачи, соответствующие их эволюционным адаптациям. Это включает в себя изменение традиционных настроек лабиринта с учетом сенсорных возможностей и подземных специализаций. Кроме того, возможно, потребуется включить нетипичные мотиваторы (например, домашнюю подстилку) и удалить конкурирующие и мешающие мотиваторы (например, туннельные тупики), чтобы оптимизировать выполнение вида в поведенческой задаче и получить истинное представление о способности пространственного обучения и памяти вида. В частности, для голых землекопов использование нагревательной лампы в сочетании с домашней подстилкой в камере вознаграждения эффективно заменило типичные мотиваторы (еда или жидкое вознаграждение после лишения пищи), которые не идеальны для животных в структуре социальной колонии, где изоляция может оказаться вредной. . Потому что H. glaber имеет много поведенческих и анатомических отличий от других батиергид, методы, используемые в текущем исследовании, вероятно, потребуют определенных модификаций, чтобы оптимизировать пространственное обучение и тестирование памяти для других представителей Bathyergidae.
. Потому что H. glaber имеет много поведенческих и анатомических отличий от других батиергид, методы, используемые в текущем исследовании, вероятно, потребуют определенных модификаций, чтобы оптимизировать пространственное обучение и тестирование памяти для других представителей Bathyergidae.
Очевидный эффект пола был очевиден в крутой кривой обучения, связанной с непрерывным тестированием лабиринта у голых землекопов, но задержка и расстояние оказались эффективными мерами пространственного обучения (рис. 5). Кроме того, приобретенное пространственное обучение сохранялось не менее 1 месяца (рис. 7). В будущих исследованиях дизайн непрерывного лабиринта, который мы разработали, выиграет от повышенной сложности, чтобы лучше проверить степень пространственного обучения и памяти. Дальнейшие временные точки (например, 2 месяца, 6 месяцев, 1 год) и сравнение показателей проб памяти с наличием мотиваторов и без них в камере вознаграждения могут помочь выяснить истинную степень сохранения пространственной памяти у голых землекопов, в дополнение к определяющие факторы, которые могут повлиять на потерю памяти.
Это исследование закладывает основу для будущих поведенческих тестов, которые будут разработаны в помощь поступательным исследованиям, таким как использование голых землекопов в нейродегенеративных исследованиях (Markey et al., 2015) или изучение нарушений пространственного обучения и памяти после потери зубов ( Okamoto et al., 2010; Chen et al., 2018; Lin et al., 2020), в дополнение к фундаментальным научным исследованиям, посвященным эволюционным адаптациям, характерным для роющих грызунов (Edrey et al., 2011; Smith et al., 2015). ; Граймс и др., 2017). Учитывая уникальную адаптацию сенсорных систем у батиергид, будущие исследования их поведенческого использования специализированных чувств, таких как магнитная ориентация (Oliveriusová et al., 2012), могут дать ценную информацию о том, как эти системы способствуют обучению и производительности памяти.
Заявление о доступности данных
Необработанные данные, подтверждающие выводы этой статьи, будут предоставлены авторами без неоправданных оговорок.
Заявление об этике
Исследование на животных было рассмотрено и одобрено Институциональным комитетом по уходу и использованию животных Университета Южного Иллинойса, Карбондейл, Иллинойс, США.
Вклад авторов
NH и DS внесли свой вклад в экспериментальный дизайн. NH и LA собрали данные для исследования. NH и KS проанализировали данные. NH, KS и DS интерпретировали данные. NH подготовил рисунки рукописи. NH и DS подготовили рукопись с исправлениями, внесенными NH, LA, KS и DS. Все авторы внесли свой вклад в статью и одобрили представленную версию.
Конфликт интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Примечание издателя
Все утверждения, изложенные в этой статье, принадлежат исключительно авторам и не обязательно представляют претензии их дочерних организаций или издателя, редакторов и рецензентов. Любой продукт, который может быть оценен в этой статье, или претензии, которые могут быть сделаны его производителем, не гарантируются и не поддерживаются издателем.
Любой продукт, который может быть оценен в этой статье, или претензии, которые могут быть сделаны его производителем, не гарантируются и не поддерживаются издателем.
Благодарности
Мы хотели бы поблагодарить Грегори Роуза за его неоценимый вклад в дизайн исследования и рецензентов за их полезные отзывы, которые значительно улучшили эту рукопись.
Дополнительный материал
Дополнительный материал к этой статье можно найти в Интернете по адресу: https://www.frontiersin.org/articles/10.3389/fevo.2022.879989/full#supplementary-material
Ссылки
D. Antinuchi, C. Antinuchi, C. и Шлейх, CE (2003). «Пространственная память у одиночного подземного грызуна Ctenomys talarum (rodentia: ctenomyidae)». Бел. Дж. Зул. 133, 89–91.
Google Scholar
Artwohl, J., Hill, T., Comer, C., и Park, T. (2002). «Голые землекопы: уникальные возможности и проблемы содержания». Лабораторное животное 31:32. doi: 10.1038/5000156
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Бароне К. М., Дума С., Рейнтьес Д.О.Дж., Броу Б.М., Кеппль К., Кламп Г. и др. (2019). «Измененная кохлеарная иннервация у развивающихся и половозрелых голых и дамараландских землекопов». J. Сравнительный анализ. Нейрол. 527, 2302–2316. doi: 10.1002/cne.24682
М., Дума С., Рейнтьес Д.О.Дж., Броу Б.М., Кеппль К., Кламп Г. и др. (2019). «Измененная кохлеарная иннервация у развивающихся и половозрелых голых и дамараландских землекопов». J. Сравнительный анализ. Нейрол. 527, 2302–2316. doi: 10.1002/cne.24682
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Bennett, NC, and Jarvis, JUM (2004). ‘Криптомис дамаренсис’. Млекопитающее. Вид 756:1. doi: 10.1644/756
CrossRef Полный текст | Google Scholar
Bennett, N.C., Faulkes, C.G., Hart, L., and Jarvis, JUM (2009). «Bathyergus suillus (rodentia: Bathyergidae)». Млекопитающее. Оксфордский университет 828, 1–7. дои: 10.1644/828.1
Полнотекстовая перекрестная ссылка | Google Scholar
Брачетта, В., Шлейх, К.Э., и Зенуто, Р.Р. (2014). «Влияние острого и хронического воздействия сигналов хищников на способности к пространственному обучению у подземных грызунов Ctenomys talarum (rodentia: ctenomyidae)». Этология 120, 563–576. doi: 10.1111/eth.12230
doi: 10.1111/eth.12230
Полный текст CrossRef | Google Scholar
Брауде С., Хесс Дж. и Ингрэм К. (2021). «Межколониальная инвазия между колониями диких голых землекопов». Дж. Зул. 313, 37–42. doi: 10.1111/jzo.12834
Полный текст CrossRef | Google Scholar
Бретт, Р. А. (1991). «Экология колоний голых землекопов: рытье нор, пища и ограничивающие факторы», в The Biology of the Naked Mole-Rat , eds PW Sherman, JUM Jarvis и RD Alexander (New Jersey: Princeton University Press), 137. –184.
Google Scholar
Браун, М. Ф., и Кук, Р. Г. (1986). «Внутрииспытательная динамика производительности лабиринта с радиальными рукавами у крыс». Учиться. Мотивировать. 17, 190–205. doi: 10.1016/0023-9690(86)-X
CrossRef Full Text | Google Scholar
Баффенштейн Р., Аморозо В., Андзиак Б., Авдеев С., Азпуруа Дж., Баркер А. Дж. и др. (2022). «Голая правда: всестороннее разъяснение и классификация современных «мифов» в биологии голого землекопа». Биол. Ред. 97, 115–140. doi: 10.1111/brv.12791
Биол. Ред. 97, 115–140. doi: 10.1111/brv.12791
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Баффенштейн Р. и Крафт В. (2021). «Идиосинкразические физиологические черты голого землекопа; устойчивая животная модель старения, долголетия и продолжительности здоровья». Доп. Эксп. Мед. биол. США 1319, 221–254. doi: 10.1007/978-3-030-65943-1_8
CrossRef Полный текст | Google Scholar
Чен Дж., Рен С. Дж., Ву Л., Ся Л. Ю., Шао Дж. и Ленг В. Д. (2018). «Потеря зубов связана с повышенным риском деменции и с зависимостью доза-реакция». Перед. Стареющие нейроски. 10:1–9. doi: 10.3389/fnagi.2018.00415
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Кларк, Ф. М., и Фолкс, К. Г. (1997). «Доминирование и смена маток в содержащихся в неволе колониях эусоциального голого землекопа, гетероцефала глабера». Проц. Р. Соц. биол. науч. 264, 993–1000. doi: 10.1098/rspb.1997.0137
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Кларк, Ф. М., и Фолкс, К. Г. (1998). Гормональные и поведенческие корреляты доминирования самцов и репродуктивного статуса в содержащихся в неволе колониях голого землекопа, Heterocephalus glaber . Биол. науч. 37, 1391–1399. doi: 10.1098/rspb.1998.0447
М., и Фолкс, К. Г. (1998). Гормональные и поведенческие корреляты доминирования самцов и репродуктивного статуса в содержащихся в неволе колониях голого землекопа, Heterocephalus glaber . Биол. науч. 37, 1391–1399. doi: 10.1098/rspb.1998.0447
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Костанцо, М.С., Беннетт, Н.К., и Лютерманн, Х. (2009). «Пространственное обучение и память у африканских землекопов: роль социальности и пола». Физиол. Поведение 96, 128–134. doi: 10.1016/j.physbeh.2008.09.008
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Криш, С. Д., Денглер-Криш, К. М., и Катания, К. К. (2006). «Центральная зрительная система голого землекопа ( Heterocephalus glaber )’. Анат. Рек. Часть А Дис. Мол. Клетка. Эвол. биол. 288, 205–212. doi: 10.1002/ar.a.20288
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Криш, С. Д., Райс, Ф. Л. , Парк, Т. Дж., и Комер, К. М. (2003). «Соматосенсорная организация и поведение голых землекопов I: волоски на теле, похожие на вибриссы, составляют сенсорный массив, который опосредует ориентацию на тактильные раздражители». Поведение мозга. Эволу. 62, 141–151. дои: 10.1159/000072723
, Парк, Т. Дж., и Комер, К. М. (2003). «Соматосенсорная организация и поведение голых землекопов I: волоски на теле, похожие на вибриссы, составляют сенсорный массив, который опосредует ориентацию на тактильные раздражители». Поведение мозга. Эволу. 62, 141–151. дои: 10.1159/000072723
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Дикон, Р. (2012). «Оценка рытья нор, строительства гнезд и накопления мышей». Дж. Визу. Эксп. 59, 1–10. doi: 10.3791/2607
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
du Toit, L., Bennett, N.C., Nickless, A., and Whiting, M.J. (2012). «Влияние пространственной среды на обучение лабиринту у африканского землекопа». Знак животного. 15, 797–806. doi: 10.1007/s10071-012-0503-0
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Эдри Ю. Х., Парк Т. Дж., Канг Х., Бини А. и Баффенштейн Р. (2011). «Эндокринная функция и нейробиология самого долгоживущего грызуна, голого землекопа». Экспл. Геронт. Эльзевир 46, 116–123. doi: 10.1016/j.exger.2010.09.005
Экспл. Геронт. Эльзевир 46, 116–123. doi: 10.1016/j.exger.2010.09.005
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Faulkes, CG, and Bennett, NC (2021). «Социальная эволюция африканских землекопов — сравнительный обзор». Доп. Эксп. Мед. биол. США 1319, 1–33. doi: 10.1007/978-3-030-65943-1_1
CrossRef Полный текст | Google Scholar
Гилберт, Дж. Д., Росситер, С. Дж., и Фолкс, К. Г. (2020). «Отношения между индивидуальным фенотипом и разделением труда у голых землекопов: это сложно». PeerJ 8:e9891. doi: 10.7717/peerj.9891
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Граймс К.М., Барефилд Д.Ю., Кумар М., Макнамара Дж.В., Вайнтрауб С.Т., де Томбе П.П. и др. (2017). «Голый землекоп демонстрирует необычный профиль белка сердечной миофиламенты, что дает новое представление о сердечной функции этого естественно подземного грызуна». Арка Пфлюгера. Евро. Дж. Физиол. Арка Пфлюгера. Евро. Дж. Физиол. 469, 1603–1613. doi: 10.1007/s00424-017-2046-3
Евро. Дж. Физиол. 469, 1603–1613. doi: 10.1007/s00424-017-2046-3
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Хеффнер Р.С. и Хеффнер Х.Е. (1993). «Дегенеративный слух и локализация звука у голых землекопов ( Heterocephalus glaber ) с обзором центральных слуховых структур». J. Сравнительный анализ. Нейрол. 331, 418–433. doi: 10.1002/cne.0311
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Генри, Э. К., Ремпл, М. С., О’Риайн, М. Дж., и Катания, К. С. (2006). «Организация соматосенсорных областей коры голого землекопа ( Heterocephalus glaber )». J. Сравнительный анализ. Нейрол. 495, 434–452. doi: 10.1002/cne.20883
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Hetling, J. R., Baig-Silva, M. S., Comer, C. M., Pardue, M. T., Samaan, D. Y., Qtaishat, N. M., et al. (2005). ‘Особенности зрительной функции голого землекопа Heterocephalus glaber’ . J. Сравнительный анализ. Физиол. Нейроэтол. Сенатор Нейронное поведение. Физиол. 191, 317–330. doi: 10.1007/s00359-004-0584-6
J. Сравнительный анализ. Физиол. Нейроэтол. Сенатор Нейронное поведение. Физиол. 191, 317–330. doi: 10.1007/s00359-004-0584-6
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Джарвис, Дж. У. М., и Беннетт, Северная Каролина (2017). «Экология и поведение семейства Bathyergidae», в The Biology of the Naked Mole-Rat , eds PW Sherman, JUM Jarvis и RD Alexander (Princeton University Press), 66–96. дои: 10.1515/9781400887132-006
Полнотекстовая перекрестная ссылка | Google Scholar
Кимчи Т. и Теркель Дж. (2001). «Пространственное обучение и память у слепыша в сравнении с лабораторной крысой и левантийской полевкой». Поведение животных. 61, 171–180. doi: 10.1006/anbe.2000.1565
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Кимчи Т. и Теркель Дж. (2004). «Сравнение роли соматосенсорных стимулов в обучении лабиринту у слепого подземного грызуна и зрячего поверхностного грызуна». Поведение. Мозг Res. 153, 389–395. doi: 10.1016/j.bbr.2003.12.015
Поведение. Мозг Res. 153, 389–395. doi: 10.1016/j.bbr.2003.12.015
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ковальзон В. М., Аверина О. А., Высоких М. Ю. (2021). «Моторная активность и «неотенический» сон голого землекопа ( Heterocephalus glaber ) в условиях изоляции». Доклады биол. науч. 496, 25–29. doi: 10.1134/S0012496621010063
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ковальзон В. М., Аверина О. А., Миньков В. А., Петрин А. А., Ю М. (2020). Необычная корреляция между ритмами отдыха-активности и температурой тела у голого землекопа ( Heterocephalus glaber ) по сравнению с пятью другими видами млекопитающих. Дж. Эвол. Биохим. Физиол. 56, 451–458. doi: 10.1134/s00220050087
CrossRef Full Text | Google Scholar
Lin, C.S., Lin, H.H., Fann, S.W., Lee, W.J., Hsu, M.L., Wang, S.J., et al. (2020). «Связь между потерей зубов и объемом серого вещества при когнитивных нарушениях». Изображение мозга. Поведение 14, 396–407. doi: 10.1007/s11682-020-00267-w
Изображение мозга. Поведение 14, 396–407. doi: 10.1007/s11682-020-00267-w
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Маркей П.М., Медина Д.Х., Гачинска М., Осмульски П.А., Оддо С., Каккамо А. и др. (2015). «Бета-амилоид и самый долгоживущий грызун: голый землекоп как модель естественной защиты от болезни Альцгеймера». Нейробиол. Старение 6, 300–308. doi: 10.1016/j.neurobiolaging.2013.03.032.Amyloid
CrossRef Полный текст | Google Scholar
Мейсон, М. Дж., Корнуолл, Х. Л., и Смит, Э. С. Дж. (2016). Ушные структуры голого землекопа, Heterocephalus glaber и его родственники (rodentia: Bathyergidae)». PLoS One 11:e0167079. doi: 10.1371/journal.pone.0167079
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Моррис, Р. (1984). «Разработка процедуры водного лабиринта для изучения пространственного обучения у крыс». J. Neurosci. Методы 11, 47–60. doi: 10.1016/0165-0270(84)-4
CrossRef Full Text | Google Scholar
Нойманн К. и Кулик Л. (2020). «Иерархии доминирования животных по рейтингу Эло», версия пакета R 0.46.
и Кулик Л. (2020). «Иерархии доминирования животных по рейтингу Эло», версия пакета R 0.46.
Google Scholar
О’Риайн, М.Дж., и Джарвис, Дж.У.М. (1997). «Распознавание членов колонии и ксенофобия у голого землекопа». Поведение животных. 53, 487–498. doi: 10.1006/anbe.1996.0299
Полный текст CrossRef | Google Scholar
Окамото Н., Морикава М., Окамото К., Хабу Н., Ивамото Дж., Томиока К. и др. (2010). «Связь потери зубов с легкими нарушениями памяти и когнитивными нарушениями: результаты исследования Фудживара-Кё». Поведение. Функция мозга. 6, 1–8. doi: 10.1186/1744-9081-6-77
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Оканоя К., Йосида С., Бароне К. М., Эпплгейт Д. Т., Бриттан-Пауэлл Э. Ф., Дулинг Р. Дж. и др. (2018). «Слухово-речевая связь у голого землекопа, млекопитающего с плохим слуховым порогом». J. Сравнительный анализ. Физиол. Нейроэтол. Сенатор Нейронное поведение. Физиол. 204, 905–914. doi: 10.1007/s00359-018-1287-8
doi: 10.1007/s00359-018-1287-8
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Оливериусова Л., Немец П., Кралова З. и Седлаек Ф. (2012). «Ориентация по магнитному компасу у двух строго подземных грызунов: приобретенное или видоспецифичное врожденное предпочтение направления?». Дж. Экспл. биол. 215, 3649–3654. doi: 10.1242/jeb.069625
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Олтон Д.С. и Самуэльсон Р.Дж. (1976). «Воспоминание о пройденных местах: пространственная память у крыс». Дж. Экспл. Психол. Поведение животных. проц. 2, 97–116. doi: 10.1037/0097-7403.2.2.97
Полный текст CrossRef | Google Scholar
Oosthuizen, MK (2020). Исследовательское поведение, память и нейрогенез у социального дамараландского землекопа ( Fukomys damarensis ). Дж. Экспл. биол. 223:1093. doi: 10.1242/jeb.221093
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Oosthuizen, M. K., Scheibler, A.G., Bennett, N.C., and Amrein, I. (2013). «Влияние лабораторного жилья на исследовательское поведение, распознавание новизны и пространственную референтную память у подземного одиночного грызуна — слепыша (9).0023 Georychus capensis )’. PLoS One 8:1–8. doi: 10.1371/journal.pone.0075863
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
R Core Team (2021 г.). R: Язык и среда для статистических вычислений. Вена: Фонд статистических вычислений R.
Google Scholar
Рюппель, Э. (1842). «Säugethiere aus der ordnung der nager, beobachtet im nordöstlichen Africa». Museum Senckenbergianum: Abhandlungen Aus Dem Gebiete der Beschreibenden Naturgeschichte 3, 99–101.
Google Scholar
Schleich, CE (2010). «Онтогенез пространственной рабочей памяти у подземного грызуна Ctenomys talarum» . Дев. Психобиол. 52, 592–597. doi: 10.1002/dev.20466
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Шерман П. В., Джарвис Дж.У.М. и Александр Р.Д. (1991). Биология голого землекопа. Принстон, Нью-Джерси: Издательство Принстонского университета.
Google Scholar
Смит, Э. С. Дж., Шумахер, Л.-Н., и Хассон, З. (2015). Голый землекоп как животная модель в медико-биологических исследованиях: современные перспективы. Открытый доступ к физиологии животных. 137:50376. doi: 10.2147/oaap.s50376
Полный текст CrossRef | Google Scholar
Смит, М., и Баффенштейн, Р. (2021). «Управляемый уход за голыми землекопами BT», в The Extraordinary Biology of the Naked Mole-Rat , eds R. Buffenstein, TJ Park и MM Holmes (Cham: Springer International Publishing), 381–407. дои: 10.1007/978-3-030-65943-1_16
CrossRef Полный текст | Google Scholar
Тур И., Клемент Д., Карлсон Е. Н. и Холмс М. М. (2015). «Обоняние и социальное познание у эусоциальных голых землекопов. Heterocephalus Glaber’, Animal Behav. 107, 175–181. doi: 10.1016/j.anbehav.2015. 06.015
CrossRef Полный текст | Google Scholar
Валлианатоу, К.А., Алонсо, А., Алеман, А., и Гензель, Л. (2020). Вызванные схемой сдвиги в навигационных стратегиях мышей раскрываются с помощью минимальной поведенческой модели пространственного исследования. bioRxiv [препринт]. doi: 10.1101/2020.12.21.423808
CrossRef Полный текст | Google Scholar
Walcher, J., Ojeda-Alonso, J., Haseleu, J., Oosthuizen, M.K., Rowe, A.H., Bennett, N.C., et al. (2018). «Специализированные механорецепторные системы в гладкой коже грызунов». J. Physiol. 596, 4995–5016.
Google Scholar
Wilson, EO (1971). Общества насекомых. Кембридж, Массачусетс: Belknap Press of Harvard University Press.
Google Scholar
Яхав С. и Баффенштейн Р. (1992). «Функция слепой кишки обеспечивает энергию брожения без выделения тепла у пойкилотермных млекопитающих Heterocephalus glaber ». J. Сравнительный анализ. Физиол. 162, 216–218. doi: 10.1007/BF00357526
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Передовая морфология зубного аппарата слепыша [PeerJ]
Введение
Змеи — очень успешные хищники, колонизировавшие многочисленные места обитания (Ганс, 19 лет).61; Грин, 1983). Их успех отчасти объясняется особой морфологией, характерным для длинного тела, отсутствия функциональных конечностей, высокой подвижности черепа и характерных зубов (Greene, 1997; Cundall & Greene, 2000; Longrich, Bhullar & Gauthier, 2012). Морфология челюстей змей приспособлена для проглатывания добычи целиком, с черепным кинезисом и несросшимися зубными костями (нижнечелюстной симфиз), обеспечивающими широкое раскрытие (Cundall & Greene, 2000), и существует разнообразный диапазон форм и расположения зубов. помогать в поимке добычи (Грин, 19 лет)97; Нокс и Джексон, 2010 г.; Vaeth, Rossman & Shoop, 1985). Однако в последние годы разнообразие зубов змей мало исследовалось, особенно для неядовитых видов, несмотря на его важность для экологии змей и его интересную изменчивость среди таксонов.
Исследования морфологии зубов змей были в основном сосредоточены на традиционных системах доставки яда (например, Fry et al., 2012; Jackson, 2003; Vaeth, Rossman & Shoop, 1985). Змеиный клык обычно описывается как длинный заостренный верхнечелюстной зуб, приспособленный для переноса и распределения яда в ткани жертвы или нападающего (Jackson, 2003). Клыки могут быть трубчатыми, с бороздками или без канавок, при этом клыки без борозд отличаются от других увеличенных зубов наличием гребней вдоль дистальной ростральной и каудальной поверхности зуба и наличием ядовитой железы (Jackson & Fritts, 19).95; Джексон, 2007). Яд мог эволюционировать, чтобы помогать захватывать и защищать добычу (Fry et al., 2006; Casewell et al., 2013) и, возможно, играть роль в облегчении пищеварения (Savitzky, 1980; Thomas & Pough, 1979), но см. McCue ( 2007 г.) для альтернативного представления. Несмотря на отсутствие трубчатых или желобчатых структур в обычных зубах, зубной ряд неядовитых видов змей не обязательно простой. Зубные гребни довольно обычны как на зубных, так и на верхнечелюстных зубах ужеобразных, таких как Dipsadinae и Natricinae, а также водных Homalopsidae, все из которых имеют разнообразный рацион и среду обитания (Bogert, 19).64; Вэт, Россман и Шуп, 1985; Янг и Кардонг, 1996).
Модификация змеиных зубов, вероятно, способствовала эволюции и распространению змей (Jackson, 2003; De Oliveira et al., 2016; Savitzky, 1980) из-за новых возможностей кормления, связанных с формой и размером добычи, которые это предлагало (Gans, 1961). ; Грин, 1983). У ужеобразных диета (а не филогенетическая история), по-видимому, действует как сильнейшее селективное давление на эволюцию, в частности, специализированного верхнечелюстного зубного ряда (Knox & Jackson, 2010). Бритт, Кларк и Беннетт (2009 г.)) постулируют, что морфология зубов у подвязочных змей напрямую связана с диетическими предпочтениями и что у кормящихся особей, как правило, имеются зубные особенности, такие как гребни, которые отсутствуют у близкородственных кормящихся универсальных змей. Верхнечелюстные зубы североамериканской западной наземной подвязочной змеи, Thamnophis elegans, , и обыкновенной волчьей змеи из Юго-Восточной Азии, Lycodon aulicus capucinus, , имеют острые гребни или кили на задних краях (Jackson & Fritts, 2004; Райт, Кардонг и Бентли, 19 лет79). Это может быть адаптация к прорезанию чешуи сцинка, способствующая глубокому проникновению зубов в добычу и предотвращающая выход добычи изо рта во время поедания. Зубы североамериканской краснобрюхой змеи Storeria occipitomaculata также имеют дистальные кили, которые помогают удерживать скользкую добычу брюхоногого моллюска (Do Amaral, 1999). Таким образом, морфология зубов неядовитых змей тесно связана с типом добычи.
Кротовая змея ( Pseudaspis cana, Linnaeus 1758) является неядовитым лампрофиидом, который при обращении с ним способен причинять болезненные укусы, иногда требующие наложения швов, причем раны более серьезные, чем обычно при укусах неядовитых змей. Структура зубов, ответственных за эти укусы, ранее не описывалась, но предполагается, что они помогают в обращении с добычей в ограниченном пространстве, защите от землекопов или самцах в бою (Alexander & Marais, 2007; Broadley, 1983). Кротовые змеи включают в свой рацион кротов и роющих грызунов (Alexander & Marais, 2007), и вполне возможно, что сложность охоты в ограниченных норах могла стимулировать эволюцию развития этого поведения укусов и соответствующей морфологии зубов (Branch, 19).98). Было зарегистрировано, что молодые кротовые змеи охотятся на сцинков (Broadley, 1983), и, таким образом, еще одной возможной функцией острых зубов является помощь в прорезании чешуи сцинков (Branch, 1998). Западная килевидная змея, Pythonodipsas carinata , родственный таксон Pseudaspis , также имеет особую морфологию зубов — считается, что ее увеличенные небные клыки помогают извлекать добычу из нор (Branch et al., 1997). Точно так же диета и условия охоты могли способствовать развитию необычного укуса кротовой змеи.
Свидетельства характерного укуса кротовой змеи также были обнаружены у самцов конспецифика в результате борьбы самцов с самцами (G Alexander, личные наблюдения, 2015). Борьба между самцами описана у многих видов змей и обычно включает ритуальную борьбу (Shaw, 1951; Carpenter, 1977; Blouin-Demers, Gibbs & Weatherhead, 2005). Змеи-кроты необычны в том отношении, что бой между самцами включает в себя укусы, которые могут привести к травмам (Greene, 1997; Branch, 1998), и раны, наносимые самцами кротових змей соперникам в виде парных параллельных порезов поперек и вокруг тела (G Александр и А. Эванс, личные наблюдения, 2015 г., рис. 1A и 1B).
Рисунок 1: Фотографии свежих ран (A) и шрамов от прошлых ран (B) на самцах слепышовых змей, полученных во время боя между самцами.
Фото: Грэм Александр (A), Николас Эванс (B). Мы изучили внешнюю структуру зубов кротовой змеи с помощью микро-КТ (компьютерной томографии) и СЭМ (сканирующей электронной микроскопии), чтобы исследовать зубы, ответственные за нанесение характерных параллельных надрезов. Мы также исследовали, присутствует ли половой диморфизм в размере, изменчивости и форме зубов в зубных рядах кротовых змей и в частоте укушенных ран на музейных экземплярах. Если кротовые змеи имеют неоднородные зубные ряды вдоль зубного ряда и демонстрируют значительные различия в форме и размере зубов между полами, то вполне вероятно, что самцы используют эти зубы для стратегии успеха в бою, особенно если параллельные следы укусов присутствуют только у самцов. . Черты, которые проявляются как у самцов, так и у самок, вероятно, больше указывают на специализацию добычи или другую адаптацию, не связанную с боем.
Материалы и методы
Головы 14 особей Pseudaspis cana (Linnaeus 1758) (7 ♂; 7 ♀), собранных в провинции Гаутенг, Южная Африка, были отсканированы с помощью микрокомпьютерной томографии (микро-КТ) в Центре микрофокусной рентгеновской КТ в Институт эволюционных наук Университета Витватерсранда, Йоханнесбург (использование образцов было одобрено Комитетом по этике исследований на животных Университета Витватерсранда, отказ № 2004/28/1). Пол каждой особи определяли с помощью зондирования клоаки. Восемь экземпляров были найдены свежеубитыми (убитыми на дороге), а остальные образцы (консервированные в этаноле или формалине) были взяты на время из Музея естественной истории Дитсонг, Гаутенг, Южная Африка. Образцы большего размера (длина головки >40 мм) сканировались при 70 кВ, 120 мкА, 1 кадр/с, 1 fa и 2000 проекций, а образцы меньшего размера (длина головы <40 мм) — при 120 кВ, 150 мкА, 1 кадр/с. , 1 фа и 2000 проекций. КТ черепа и зубного аппарата каждого образца были реконструированы с помощью CT Pro 3D, а затем сегментированы в цифровом виде и проанализированы с использованием программного обеспечения для визуализации КТ VG Studio Max 3.0 (Volume Graphics, 2014). Это программное обеспечение позволило создать точные 3D-модели с высоким разрешением зубного аппарата змеи и его соединения с черепным скелетом.
Зубы Dendentary были исследованы путем подсчета количества зубов с острыми краями и сравнения между отдельными особями и полами. Заметные различия в размерах зубов верхней челюсти в разных положениях вдоль челюсти (в отличие от довольно одинакового размера зубов нижней челюсти) побудили нас измерить и сравнить объемы зубов верхней челюсти. Зубы верхней челюсти анализировали путем сравнения положения, формы и размера зубов в различных положениях на верхней челюсти у каждого человека. Объемы были извлечены в VG Studio Max 3.0 после цифровой сегментации отдельных зубов и челюстей черепа. Длина зубьев измерялась с помощью Avizo 9.но не использовались в анализе, потому что различия в степени кривизны зубов (которые не измерялись из-за высоких различий внутри и между отдельными людьми) означали, что длины не давали надежного измерения размера зубов.
Были проведены измерения зубов верхней челюсти для двух самых передних зубов (T1 и T2), двух зубов посередине верхней челюсти (T6 и T7) и трех самых задних зубов (T11–T13) как левой, так и правой верхней челюсти, чтобы исследовать вариация зубов вдоль верхней челюсти (рис. 2). Значения для недоразвитых зубов (еще не прикрепленных к зубочелюстному элементу) были исключены из анализа. Объем зуба измеряли как общий объем зуба с цифровым сегментированием. Объемы нижней правой и левой челюстей (составной и зубной костей вместе) были измерены и использованы в качестве косвенного показателя размера черепа и средства оценки эффектов аллометрии из-за довольно жесткой структуры нижней челюсти. Соединение между зубной и сложной костями заполнено коллагеновой тканью, которая может допускать небольшое количество движений, но это очень ограничено по сравнению с движением и, следовательно, аддитивной ошибкой, которая была бы связана с измерением всего черепа с несколькими кинетические части.
Рисунок 2: Микро-КТ (компьютерная томография) реконструкция левой верхней челюсти слепыша, показывающая зубы верхней челюсти, выбранные для статистического анализа.
Частота укусов кротовых змей была исследована на 50 образцах из Музея естественной истории Дитсонг. Каждую змею осматривали на наличие открытых ран или шрамов и отмечали пол. Мы записали все порезы на теле и отметили серьезность. Хотя не всегда возможно окончательно отличить раны, полученные в результате единоборства, от ранений, полученных по другим причинам, предполагается, что поперечные порезы более 25% ширины тела, в частности спиральные порезы, вероятно, были нанесены в бою между самцами. , в то время как чешуя или очень короткие шрамы (<25% ширины спинной поверхности) не считались результатом укусов сородичей. Положение разрезов регистрировали либо в области хвоста (от клоаки до кончика хвоста), либо в области туловища каждого экземпляра.
Деталь зуба была качественно исследована с помощью сканирующей электронной микроскопии (СЭМ). Зубы, представляющие интерес (при исследовании 3D-моделей КТ), были покрыты 10-нм углеродом и 5-нм сплавом золото-палладий (60:40), а затем были микрофотографированы с использованием FEI Quanta 200 E-SEM в Отделе микроскопии и микроанализа в в школе наук о животных, растениях и окружающей среде Университета Витватерсранда в Йоханнесбурге.
Анализ данных
Все статистические анализы проводились с использованием RStudio версии 3.2.5 (R Core Team, 2014 г.). Измерения объема зубов использовались в серии сравнений, зуб за зубом, внутри отдельных людей, между людьми и между полами. Чтобы учесть различия в размерах отдельных змей, объемы зубов каждой особи сначала делили на объем нижней челюсти соответствующей особи. Среднее значение левого и правого зубов для каждого из измеряемых зубов рассчитывали для каждого образца (например, объемы Т1 на левой верхней челюсти и Т1 на правой верхней челюсти, чтобы получить средний объем Т1 для образца). То же самое было сделано для измерений нижней левой и правой челюсти, чтобы получить средний объем нижней челюсти.
После проверки на нормальность с помощью теста Шапиро-Уилка односторонний ANOVA был использован для измерения внутрииндивидуальной вариации логарифмически преобразованных объемов зубов в различных положениях на верхней челюсти. Половые различия были проверены с использованием многофакторного дисперсионного анализа с учетом пола в качестве фактора. Тесты проводились с семью средними объемами зубов верхней челюсти в положениях, описанных выше (T1, T2, T6, T7, T11, T12 и T13; см. рис. 2). Критерии суммы рангов Уилкоксона использовались для проверки того, различаются ли объемы нижних челюстей, количество зубных зубов и средний объем зубов верхней челюсти у самцов и самок слепышовых змей. Тест хи-квадрат проверял, существует ли связь между частотой ран от укусов сородичей и полом.
Результаты
Морфология верхнечелюстных и нижнечелюстных зубов слепыша
Каждая верхняя и зубная кости имели непрерывный ряд твердых акродонтных зубов (прикрепленных непосредственно к кости челюсти), которые различались по размеру и форме (рис. 2 и 3). Сменные зубы (удаленные с КТ-изображений) лежат с медиальной стороны функциональных зубов.
Рисунок 3: Микро-КТ (компьютерная томография) реконструкция левого бокового вида черепа самца кротовой змеи, показывающая череп (cr), квадратную (q), составную (co), зубную (d) и верхнюю челюсть (m) кости.
Зубной ряд
Каждая из отдельных зубных костей имела от 14 до 16 зубов. Эти зубы были загнуты назад, а некоторые имели задние кили (острые гребни/края) (рис. 3 и 4), в то время как зубы без острых краев были просто коническими, сужающимися к острию. Количество зубов с острыми килями варьировало у разных змей, от 5 до 13 на каждой зубной кости, и отсутствовало у некоторых экземпляров (см. Половые различия относительной формы и объема зубов 9).0024 ниже). Когда они присутствовали, кили встречались в самых задних зубах.
Рисунок 4: СЭМ (сканирующая электронная микроскопия) изображение заднего нижнечелюстного зуба самца слепыша, на заднем крае которого виден лезвиеобразный киль.
Зубы верхней челюсти
Зубы верхней челюсти различались по размеру и форме вдоль каждой кости верхней челюсти с заметными различиями между передними и задними зубами (рис. 2). Количество зубов верхней челюсти было в основном одинаковым (13 зубов с каждой стороны) у всех образцов, при этом у одного образца в этом исследовании было 14 зубов на одной верхней челюсти. Зубы на переднем конце и в середине верхней челюсти были длинными, коническими, загнутыми в основании зубов (рис. 2). Два самых задних зуба (T12 и T13) имели характерную форму: они были лабио-лингвально уплощенными и треугольными, а не коническими, а в некоторых образцах были направлены почти прямо кзади от основания зуба, а не перпендикулярно и слегка загибались назад к опорному зубу. maxilla, как и в случае более передних зубов (рис. 2 и 5). Т12 и Т13 также имели острые задние/дорсальные края у всех экземпляров (рис. 5).
Рисунок 5: СЭМ (сканирующая электронная микроскопия) изображения задних зубов верхней челюсти (T12, T13) самца слепыша (A), подчеркивающие лезвиеобразный киль вдоль заднего края T13 (B).
ANOVA-тест объемов T1, T2, T6, T7, T11, T12 и T13 для всей выборки слепышей показал, что зубы, расположенные вдоль верхней челюсти, значительно различаются по объему по сравнению друг с другом ( F (1,6) = 12,37, р <0,001; Таблица 1). Апостериорный тест Тьюки показал, что зубы на заднем конце верхней челюсти (T12 и T13) были значительно больше, чем зубы T1, T2, T6, T7 и T11 (таблица 1). Средний объем Т13 более чем в два раза превышал объем Т1, Т2, Т6, Т7 и Т11 ( p <0,01 для всех сравнений), а Т12 был как минимум на 40% больше, чем Т1 ( p <0,001), Т2 ( p <0,01) и T11( p <0,001) (рис. 6; табл. 1).
Значимые результаты тестов ANOVA и post hoc , сравнивающих объемы зубов в различных положениях на верхней челюсти для всей выборки (14) и для отдельных полов (7m, 7f) слепышей.
| Результат проверки гипотезы | Попарное сравнение зубов | Весь образец | Кобели | Самки |
|---|---|---|---|---|
| Дисперсионный анализ p -значение | <0,000001 | 0,000029 | 0,000089 | |
| ANOVA F статистика | 12. 370000 | 7.153000 | 6.2 | |
| Tukey post hoc p -значение | Т12-Т1 | 0,000093 | 0,001468 | |
| Т13-Т1 | <0,000001 | 0,000568 | 0,001129 | |
| Т12-Т11 | 0,000015 | 0,009328 | 0,005811 | |
| Т13-Т11 | 0,000001 | 0,000023 | 0,004531 | |
| Т2-Т12 | 0,009367 | 0,028693 | ||
| Т2-Т13 | 0,000042 | 0,006182 | 0,022934 | |
| Т6-Т13 | 0,000447 | 0,002442 | ||
| Т7-Т13 | 0,001529 | 0,017644 |
DOI: 10. 7717/peerj.6943/таблица-1
Рисунок 6: Средние объемы (мм
3 ) и стандартные ошибки передних (T1 и T2), средних (T6 и T7) и задних (T11–T13) зубов верхней челюсти семи самцов слепыша (M) и семи самок слепыша. (Ф).Половые различия формы и объема зубов
Наиболее выраженные половые различия в зубном ряду были в зубных рядах. Все самцы имели задние кили (острые края) от 5 до 13 зубов на каждой зубной кости. У самок отсутствовали острые задние края на нижнечелюстных зубах, их зубы имели закругленные края по сравнению с отчетливо острыми зубами у самцов. У каждого экземпляра было от 14 до 16 зубов на каждой зубной кости, но вариации числа зубов не зависели от пола (критерий суммы рангов Уилкоксона, 9).0023 Вт = 27,5, p >0,9).
В среднем зубы, расположенные вдоль кости верхней челюсти, значительно различались по объему по сравнению друг с другом как для мужчин, так и для женщин (однофакторный дисперсионный анализ для каждого пола, мужчины: F (1,6) = 7,153, p <0,001; самки: Ж (1,6) = 6,292, p <0,001; табл. 1). Апостериорный тест Тьюки показал, что у мужчин среднее значение T13 было больше, чем T1 ( p <0,001), T2 ( p <0,01), T6 ( p <0,01), T7 ( p = 0,02) и T11 ( p <0,001), а T12 больше, чем T11 ( p <0,01) (табл. 1). У самок Т13 был больше, чем Т1 ( p <0,01), Т2 ( p = 0,02) и Т11 ( p <0,01), а Т12 был аналогично больше, чем Т1 ( p <0,01), Т2 ( p = 0,03) и T11 ( p <0,01) (рис. 6; табл. 1).
Четкие формы и большие размеры Т12 и Т13 по сравнению с другими зубами у самцов проявлялись более ярко, чем у самок (рис. 6 и 7). Критерий суммы рангов Уилкоксона не смог выявить значимой разницы между полами для среднего объема T12 ( W = 24, p > 0,9) и T13 ( W = 36, p = 0,165). Однако многофакторный дисперсионный анализ с числом зубов, полом и взаимодействием пола и зуба в качестве факторов выявил влияние пола на объем зубов (9).0023 F (1,1) = 5,512, p = 0,0213), что указывает на то, что пол способствовал различиям в средних объемах зубов между людьми (видно на рис. 6). Не было никакой разницы между средними объемами нижней челюсти мужчин и женщин (мужчины 46,9 ± 6,3 мм 3 , женщины 44,5 ± 5,1 мм 3 ; W = 23, p = 0,90), что указывает на то, что различия в относительные размеры зубов верхней челюсти не были артефактами различий в объемах челюстей.
Рисунок 7: Реконструкции микро-КТ (компьютерной томографии), сравнивающие боковую проекцию левой верхней челюсти, зубной кости и сложных костей самки (A) с таковой самца (B) слепыша.
Шрамы на музейных экземплярах
Доля самцов со шрамами или ранами была значительно выше, чем доля самок со шрамами или ранами ( х 2 = 5,5987, d , f . = 1, p ,1 8 = ). Из 50 изученных музейных экземпляров (28 взрослых самцов, 14 взрослых самок и 8 ювенильных) 9у взрослых самцов и у 1 взрослой самки были обнаружены следы рубцов или ран различной длины и глубины по оси тела. Шесть из этих самцов имели шрамы в области хвоста, начиная от рубца, огибающего хвост дважды по спирали у двух экземпляров (например, на рис. 1А), до пореза или рубца на вентральной поверхности хвоста у трех экземпляров и до небольшого , неглубокий рубец только на части дорсальной поверхности у одного экземпляра. У этого экземпляра также были параллельные надрезы, проникающие через брюшную чешую в мышцу ближе к середине туловища. Рубцы остальных трех самцов были обнаружены ближе к середине змеи и также различались по степени тяжести: от одного длинного рубца на брюшной поверхности тела до множества мелких рубцов, не покрывающих ширину змеи как на спинной, так и на брюшной поверхностях. тела. У единственной самки был неглубокий рубец над хвостом, который в этом месте составлял половину ширины спинной поверхности.
Обсуждение
Способность кротовой змеи наносить режущие укусы была отмечена рядом полевых проводников и неосторожных дрессировщиков, но до этого исследования не сообщалось о том, какие зубы вызывают это, и различаются ли эти зубы у разных полов. Явный признак полового диморфизма очевиден в зубных зубах кротовой змеи, некоторые из которых имеют заметно острые задние края (кили) только у самцов. У обоих полов два самых задних зуба на верхней челюсти заметно крупнее, лабио-лингвально уплощены, а их апикобазальная ось сильно ориентирована назад, тогда как у более передних зубов верхней челюсти она ориентирована дорсовентрально. Хотя эти верхнечелюстные зубные признаки присутствуют у обоих полов, что указывает на возможную неспецифическую для пола функцию, задние верхнечелюстные зубы более выражены у мужчин. Половой диморфизм зубов, а также более высокая частота рваных ран у самцов указывают на то, что у самцов слепышовых змей есть специальные зубные ряды, которые помогают в бою.
Вполне вероятно, что острые нижние зубы, а также задние верхнечелюстные зубы играют важную роль в создании режущих ран, типичных для укуса слепыша. Когда пасть полностью открыта, задние верхнечелюстные зубы (T12 и T13) направлены вниз и, вероятно, будут первыми зубами, вступающими в контакт с целью, как у некоторых других змей (Wright, Kardong & Bentley, 1979). Поведенческие исследования необходимы, чтобы установить точное движение челюстей и зубов во время укуса, но мы предполагаем, что есть два способа, которыми кротовая змея может укусить, чтобы вызвать эти раны: задние верхнечелюстные зубы закрепляют цель, в то время как нижняя челюсть, благодаря двойному сочленению квадратной кости он отклоняется назад, позволяя нижнечелюстным зубам прорезать добычу, гипотеза, которая согласуется с наличием более серьезных ран на вентральных поверхностях музейных образцов или задних верхнечелюстных зубах. вызывают раны, когда верхняя челюсть впервые соприкасается с соперничающей змеей во время укуса.
Рваные раны, подобные тем, которые вызваны укусами слепышовых змей, не были задокументированы у многих неядовитых видов змей, и единственными сопоставимыми видами являются австралийский ковровый питон ( Morelia spilota ) и змея кукри ( Oligodon formosanus ), оба вида которые участвовали в агрессивных боях сородичей (Shine & Fitzgerald, 1995; Huang et al. , 2011). Как и кротовая змея, ковровый питон наносит серьезные, похожие на раны укусы соперничающих самцов, соревнуясь за самку, но область тела, на которую направлены укусы, не задокументирована. Увеличенные верхнечелюстные зубы змеи кукри служат для нанесения режущих ран соперникам (неспецифическим по полу) при защите кормовой территории, а также помогают вскрывать черепашьи яйца (Huang et al., 2011). Самцы змей кукри чаще кусают хвост, чем самки, но укусы самцов змей кукри, как правило, менее серьезны, чем у самок. Хуанг и др. (2011) предполагают, что это развилось в ответ на риск повреждения репродуктивных органов.
В отличие от змеи кукри, у самцов кротовых змей больше шансов повредить хвост (и, в некоторой степени, туловище), чем у самок. Наличие задних килей на зубных зубах самцов слепыша и отсутствие этого признака у самок, а также более заметно увеличенные задние верхнечелюстные зубы самца слепыша позволяют предположить, что эти признаки могут давать некоторое преимущество, например, во время драки самцов наблюдаются в период размножения (Alexander & Marais, 2007; Broadley, 19 лет). 83). С точки зрения адаптации преимущество наличия зубов, которые наносят длинные или спиральные порезы в бою, а не более распространенные колотые раны, может заключаться в том, что они наносят больший ущерб хвосту конкурирующего самца, где расположены гемипены, уменьшая шансы соперника на поражение. репродуктивный успех, чтобы увеличить количество самок, доступных для победителя. Заметим, однако, что мы не можем сделать вывод о том, что самки не наносят укусов самцам, а также что хвост может быть просто наиболее доступной частью анатомии самца во время боя (Huang et al., 2011). Шанс повредить репродуктивные органы соперника, даже если он не нацелен конкретно на этот регион, мог привести к тому, что это поведение стало избранным в популяции.
Наличие специализированных верхнечелюстных зубов у обоих полов слепыша предполагает, что эти зубы либо обеспечивают одинаковые преимущества для каждого пола, либо являются результатом вынужденной эволюции у самок ( sensu Gould & Lewontin, 1979; Gould, 1993). Специализированные зубы верхней челюсти довольно часто встречаются у колуброидов (Vaeth, Rossman & Shoop, 1985; Scanlon & Shine, 1988; Jackson & Fritts, 1995) и, как правило, коррелируют со специализацией в питании (Knox & Jackson, 2010). Наиболее вероятное адаптационистское объяснение наличия острых треугольных задних зубов верхней челюсти у обоих полов слепыша состоит в том, что эти зубы связаны с особыми потребностями в питании. Увеличенные задние верхнечелюстные зубы (а у некоторых видов и крыловидные зубы) обычно связаны с поеданием скользкой добычи, такой как рыба (Wright, Kardong & Bentley, 19).79) или оофагии (Scanlon & Shine, 1988). Несмотря на то, что рацион кротовых змей недостаточно задокументирован (взрослые особи питаются в основном мелкими роющими млекопитающими, а молодые особи преимущественно ящерицами, Alexander & Marais, 2007; состоит ли она из особо скользкой добычи. Тем не менее, Бродли (1983) и Дайер (1996) сообщают о том, что слепыши поедают целые птичьи яйца, которые могут быть важным компонентом рациона в некоторых популяциях. Таким образом, гипотеза адаптационистской диеты может быть частичным объяснением специализированных зубов верхней челюсти у кротовых змей.
Родственный таксон кротовой змеи, западная килевидная змея ( Pythonodipsas carinata ), имеет увеличенные небные «клыки», которые могут помочь в извлечении добычи из нор (Branch et al., 1997) или для удерживания гекконов (Marx et al., 1997). др., 1982). Точно так же среда обитания для охоты, а не анатомия добычи, возможно, способствовала выбору специализированных зубов у кротовой змеи, если зубы помогали добывать добычу в ограниченных роющих пространствах, в которых кротовые змеи иногда охотятся. Однако в литературе это мало подтверждается, и, в отличие от слепыша, неядовитые роющие змеи, как правило, имеют маленькие, неизменные зубы (Савицки, 19).83).
Альтернативное объяснение специализации зубов верхней челюсти у кротовых змей состоит в том, что задние зубы являются филогенетическими остатками клыков предков. Джексон (2007) утверждает, что исходным состоянием Colubroidea было наличие трубчатых клыков на верхней челюсти, и что неядовитые виды в этой кладе, следовательно, должны были впоследствии утратить клыки и способность производить яд. Кроме того, она отличает клыки без бороздок от увеличенных зубов по наличию килей вдоль ростральной и каудальной поверхностей зуба — признаков, которые присутствуют в увеличенных верхнечелюстных зубах у кротов. Согласно Кардонгу (1982), мы не определяем зубы слепыша как клыки из-за отсутствия ядовитых желез и исключительно мукоидной функции желез Дювернуа (Taub, 1967). Однако наличие этих зубов у кротовых змей может указывать на происхождение клыков предков без канавок, которые были использованы в качестве точки привязки, позволяя нижней челюсти разрезать тела соперников-самцов, сохраняя при этом чужие (посредством отсутствие отбора для потери зубов), или для выгоды при отправке добычи или поедании яиц у самок.
Выводы
Наши наблюдения проливают свет на малоизученную кротовую змею и могут послужить основой для будущих функциональных исследований змеиных зубов, захвата добычи и полового диморфизма. Дальнейшее исследование могло бы выяснить, склонны ли самцы кротовых змей кусать хвосты соперников больше, чем остальную часть тела, и как повреждение хвоста влияет на репродуктивный успех кротовых змей. Изучение охотничьего поведения и кинетики черепа может установить точный механизм укуса слепыша, а функциональное сравнение с другими видами будет способствовать более полной картине характерного зубного аппарата слепыша.
Благодарности
Особая благодарность Кудакваше Джакате и Жаку Герберу за обучение и руководство в области компьютерной томографии и электронной электронной микроскопии. Большое спасибо Лауретте Махлангу из Музея естественной истории Дитсонга, Филипу Джордану, Адриану Стейну и Клинту Халкетт-Сиддаллу за предоставленные образцы, а также Майку Перри за то, что он позволил нам сфотографировать его живых кротовых змей.
Ссылки
Александр Г., Марэ Ж. 2007 .
Путеводитель по рептилиям юга Африки. Кейптаун: Струик.Блуэн-Демерс Г., Гиббс Х.Л., Уэзерхед П.Дж. 2005 . Генетические доказательства полового отбора у черных крысиных змей, Elaphe obsoleta . Поведение животных 69 :225-234
Богерт СМ. 1964 . Змеи родов Diaphorolepis и Synophis и ужеобразные подсемейства Xenoderminae. Senckenbergiana Biologica 45 :509-531
Отделение WR. 1998 . Полевой определитель змей и других рептилий юга Африки. Кейптаун: Струик.
Бранч В., Шайн Р., Харлоу П., Уэбб Дж. 1997 . Половой диморфизм, рацион и аспекты размножения западной килевидной змеи, Pythonodipsas carinata (Serpentes: Colubridae) African Journal of Herpetology 46 :89-97
Бритт Э.
, Кларк А., Беннетт А. 2009 . Морфология зубов подвязочных змей ( Thamnophis ) и их связь с диетическими предпочтениями. Журнал герпетологии 43 :252-259Бродли Д.Г. 1983 . Змеи Фиц-Саймонса из Южной Африки. Йоханнесбург: Книги Дельты.
Плотник CC. 1977 . Общение и проявления змей. Американский зоолог 17 :217-223
Кейсвелл Н.Р., Вюстер В., Вонк Ф.Дж., Харрисон Р.А., Фрай Б.Г. 2013 . Сложные коктейли: эволюционная новизна ядов. Тенденции в области экологии и эволюции 28 :219-229
Кандалл Д., Грин Х.В. 2000 .
Кормление змей. В кн.: Питание: форма, функции и эволюция четвероногих позвоночных. Сан-Диего: Академическая пресса. 293-333Де Оливейра Л., Скартоццони Р.Р., Де Алмейда-Сантос С.М., Джаред С., Антониацци М.М., Да Саломао Г.М. 2016 . Морфология желез Дювернуа и зубов верхней челюсти и возможная функция секреции желез Дювернуа у Helicops Modetus Günther, 1861 (Serpentes: Xenodontinae) South American Journal of Herpetology 11 :54-65
У Амарала JPS. 1999 . Скручивание губ у краснобрюхих змей ( Storeria occipitomaculata ): функциональная морфология и экологическое значение. Журнал зоологии 248 :289-293
Красильщик БМ. 1996 . Нападение змей на морских птиц на трех южноафриканских островах.
South African Journal of Marine Science 17 :309-313Фрай Б.Г., Кейсуэлл Н.Р., Вюстер В., Видал Н., Янг Б., Джексон Т.Н. 2012 . Структурная и функциональная диверсификация ядовитой системы рептилий Toxicofera. Токсикон 60 :434-448
Фрай Б.Г., Видал Н., Норман Дж.А., Вонк Ф.Дж., Шейб Х., Рамджан С.Р., Куруппу С., Фунг К., Хеджес С.Б., Ричардсон М.К. 2006 . Ранняя эволюция ядовитой системы ящериц и змей. Природа 439 :584-588
Ганс К. 1961 . Механизм питания змей и его возможная эволюция. Американский зоолог 1 :217-227
Гулд С.Дж. 1993 .
Мужские соски и рябь клитора. Колумбия: журнал литературы и искусства 20 :80-96Гулд С.Дж., Левонтин Р.К. 1979 . Петли Сан-Марко и панглоссианская парадигма: критика адаптационистской программы. Труды Лондонского королевского общества. Серия B: Биологические науки 205 :581-598
Грин ХВ. 1983 . Диетические корреляты происхождения и излучения змей. Американский зоолог 23 :431-441
Грин ХВ. 1997 . Змеи: эволюция тайны в природе. Беркли: Калифорнийский университет Press.
Хуанг В.С., Грин Х.В., Чанг Т.Дж., Шайн Р. 2011 . Территориальное поведение тайваньских кукризнейков ( Oligodon formosanus ) Proceedings of the National Academy of Sciences of the United States of America 108 :7455-7459
Джексон К.
2003 . Эволюция систем доставки яда у змей. Зоологический журнал Линнеевского общества 137 :337-354Джексон К. 2007 . Эволюция ядовитых клыков: выводы из биологии развития. Токсикон 49 :975-981
Джексон К., Фриттс Т.Х. 1995 . Доказательства морфологии поверхности зуба для заднего верхнечелюстного происхождения клыка протероглифа. Амфибии-Рептилии 16 :273-288
Джексон К., Фриттс Т.Х. 2004 . Специализация зубов при дурофагии у обыкновенной волчьей змеи, Lycodon aulicus capucinus . Амфибия-Рептилия 25 :247-254
Кардонг К.
В. 1982 . Эволюция ядовитого аппарата змей от ужеобразных до гадюк и элапидов. Memrias do Instituto Butantan 46 :105-118Нокс А., Джексон К. 2010 . Экологические и филогенетические влияния на зубной ряд верхней челюсти у змей. Phyllomedusa: журнал герпетологии 9 :121-131
Longrich NR, Bhullar B-AS, Gauthier JA. 2012 . Переходная змея из позднемелового периода Северной Америки. Природа 488 :205-208
Маркс Х., Рабб Г.Б., Арнольд С.Дж., Арнольд С.Дж. 1982 . Pythonodipsas и Spalerosophis , роды ужеобразных змей, сходные с гадюками. Копейя 3 :553-561
МакКью, Мэриленд.
2007 . Отравление жертвы не улучшает пищеварительную функцию западных гремучих змей (Crotalus atrox)R Основная команда. 2014 . R: язык и среда для статистических вычислений. Вена: Фонд статистических вычислений R. программное обеспечение
Савицкий А.Х. 1980 . Роль стратегии доставки яда в эволюции змей. Эволюция 34 :1194-1204
Савицкий А.Х. 1983 . Коадаптированные комплексы признаков у змей: окаменелость, рыбоядность и дурофагия. Американский зоолог 23 :397-409
Скэнлон Дж.Д., Шайн Р. 1988 . Зубной ряд и диета у змей: приспособление к оофагии у австралийского рода элапидов Симоселэпс .
Журнал зоологии 216 :519-528Shaw CE. 1951 . Бой самцов американских ужеобразных змей с примечаниями к боевым действиям других ужеобразных и неуклюжих змей. Herpetologica 7 :149-168
Шайн Р., Фицджеральд М. 1995 . Различия в системах спаривания и половой диморфизм между популяциями австралийского питона Morelia spilota (Serpentes: Pythonidae) Oecologia 103 :490-498
Тауб А.М. 1967 . Сравнительные гистологические исследования железы Дювернуа ужеобразных змей. Бюллетень AMNH 138 :статья 1
Томас Р., Паф Ф.Х. 1979 .
Влияние яда гремучей змеи на переваривание добычи. Токсикон 17 :221-228Vaeth RH, Rossman DA, Shoop W. 1985 . Наблюдения за морфологией поверхности зубов змей. Журнал герпетологии 19 :20-26
Объемная графика. 2014 . VGStudio Макс. программное обеспечение
Райт Д.Л., Кардонг К.В., Бентли Д.Л. 1979 . Функциональная анатомия зубов западной наземной подвязочной змеи, Thamnophis elegans . Герпетология 35 :223-228
Янг Б.А., Кардонг К.В. 1996 . Элементы поверхности зубов у змей (Reptilia: Serpentes) Amphibia-Reptilia 17 :261-276
Мотокультиватор Крот
Содержимое
- 1
Обзор популярных моделей
- 1. 1
Мотокультиватор МК-1А
- 1.2 Мотокультиватор Крот 2 с реверсом
- 1.
- 2 Руководство по эксплуатации мотокультиватора Крот
- 3 Модернизация модели МК-1А
Производство отечественных мотокультиваторов марки «Крот» было налажено в конце 80-х годов. Первая модель МК-1А оснащалась двухтактным бензиновым двигателем объемом 2,6 л. из. Запуск осуществлялся от тросового ручного стартера. Изначально техника предназначалась для обработки небольших огородов на даче и работы внутри теплицы. Современный мотокультиватор Крот представляет усовершенствованную модель МК-1А. Эта техника уже оснащена мощным двигателем с принудительным воздушным охлаждением.
Обзор популярных моделей
Примерные размеры оборудования находятся в пределах:
- длина — от 100 до 130 см;
- ширина — от 35 до 81 см;
- рост — от 71 до 106 см.
Габариты культиватора Крот зависят от модели, и могут меняться по мере совершенствования техники.
Мотокультиватор МК-1А
Начнем обзор культиваторов «Крот» с модели МК-1А. Агрегат оснащен двухтактным карбюраторным двигателем мощностью 2,6 л.с. В качестве пускового устройства используется тросовая рукоятка. Бензиновый двигатель с коробкой передач имеет простое болтовое соединение с рамой. Топливный бак рассчитан на 1,8 литра. Такой небольшой объем обусловлен малым расходом топлива. Агрегат можно заправлять дешевым бензином АИ-80 или А-76. Для приготовления топливной смеси используется машинное масло марки М-12ТП. Культиватор весит всего 48 кг. Такое оборудование легко перевезти на дачу на автомобиле.
На рукоятках расположены все элементы управления мотокультиватора, а именно:
- рычаг сцепления;
- рычаг управления дроссельной заслонкой;
- Рычаг управления заслонкой карбюратора.
Модель «Крот МК-1А» способна работать с навесным оборудованием. Мотокультиватор используется для полива, скашивания травы, обработки почвы и ухода за насаждениями.
Мотокультиватор Крот 2 с реверсом
Особенностью конструкции является то, что культиватор «Крот» имеет реверс и мощный двигатель. Это дает возможность потребителю за небольшие деньги получить настоящий мотоблок. Агрегат приводится в движение четырехтактным бензиновым двигателем Honda GX200 объемом 6,5 л. из. Крот 2 имеет электронное зажигание, вал отбора мощности, бензобак на 3,6 литра. Крутящий момент от двигателя к шасси передается ременной передачей.
Среди других мотоциклов с аналогичными характеристиками эта модель Крота занимает первые позиции по надежности. Эти показатели были достигнуты благодаря мощному одноцилиндровому мотору и надежной коробке передач. Ресурс двигателя 3500 часов. Это довольно много по сравнению со старыми моделями культиватора «Крот», имевшими моторесурс до 400 часов.
Важный! Большой плюс четырехтактного двигателя в том, что масло и бензин хранятся отдельно. Владельцу больше не нужно вручную готовить топливную смесь, смешивая эти компоненты.
Мощности мотокультиватора с задним ходом хватает, чтобы фрезы захватили участок шириной 1 м. В инструкции по эксплуатации от завода производителя сказано, что мотокультиватор Крот 2 способен расширить свой функционал за счет использования навесного оборудования. Так, техника может стать снегоуборщиком или косилкой, транспортным средством для перевозки грузов, машиной для выполнения многих сельскохозяйственных работ.
Важный! Рукоятки мотокультиватора Крот 2 имеют многоступенчатую регулировку. Оператор может поворачивать их в любую сторону, что позволяет оптимально настроить оборудование под любой вид работ.
В видео предлагаем посмотреть обзор культиватора Крот:
Руководство по эксплуатации мотокультиватора Крот
Итак, мы выяснили, что современный культиватор «Крот» имеет практически все функции мотоблока. А теперь посмотрим, что написано в инструкции по эксплуатации рассматриваемой техники:
- Прямое назначение культиватора – вспашка земли. Делается это с помощью фрез, которые устанавливаются на валы редуктора. Транспортные колеса поднимаются во время вспашки. Сошник прикреплен к задней части прицепной скобы. Используется в качестве тормоза, а также для регулировки глубины обработки почвы. Культиватор движется за счет вращения фрез, одновременно разрыхляя почву. Устройство поставляется с двумя внутренними и внешними резаками. Первый тип используется на неровной почве и целине. Легкий грунт рыхлят обоими фрезами, можно добавить и третий комплект. Купите его отдельно. В итоге резцов по три с каждой стороны, а всего их 6 штук. На культиватор «Крот» нельзя поставить восемь фрез из-за повышенной нагрузки на мотор и трансмиссию.
- При прополке сорняков проводят переоборудование механизма. На внутренние фрезы снимаются ножи, а на их место ставятся прополочные. Эти детали узнаваемы по L-образной форме. Внешние фрезы заменены дисками. Они также продаются отдельно. Диски нужны для защиты растений, не допуская их попадания под прополочную машину. Если на картофеле проводится прополка, то одновременно можно проводить и предварительное окучивание. Для этого задний сошник заменяют окучником.
- Когда нужно окучить картошку, фрезы не нужны. Их снимают с вала редуктора, а на это место ставят стальные колеса с приваренными проушинами. Румпель остается на месте, где раньше был сошник.
- При уборке картофеля используются те же металлические проушины, а за культиватором сошник заменен на картофелекопалку. Этот вид навесного оборудования выпускается в различных модификациях, но обычно для культиваторов покупают веерные модели.
- Вспахивать землю можно не только фрезами, но и плугом. Он крепится к задней части машины вместо сошника. Стальные колеса остаются на месте.
- Агрегат можно использовать на сенокосе. Нужно просто купить косилку и закрепить ее перед агрегатом. На валы редуктора надеваются резиновые колеса. Передача крутящего момента обеспечивается ремнями, надетыми на шкивы культиватора «Крот» и косилки.
- Крот вполне способен заменить насос для откачки воды. Вам достаточно приобрести насосное оборудование МНУ-2, закрепить его на раме и соединить с ременной передачей. При этом важно не забыть снять ремень с тягового механизма.
- Мотокультиватор хорошо справляется с перевозкой малогабаритных грузов массой до 200 кг. Здесь нужна тележка с поворотно-сцепным механизмом. Можно приобрести заводскую модель ТМ-200 или сварить ее самостоятельно из металла. Во время перевозки грузов на валы редуктора надеваются резиновые колеса.
Как видите, благодаря дополнительному оборудованию многофункциональность Крота значительно расширена.
Модернизация МК-1А модель
Если у вас есть старая модель «Крота», не спешите ее выбрасывать. Зачем тогда переплачивать при покупке нового культиватора за раму, редуктор и другие детали, если они уже есть. Можно обойтись простой заменой мотора.
Старый двигатель можно заменить на четырехтактный LIFAN — 160F.