Вышка-тура ПСРВ » Компания «ВНК»
| Модель | Общая высота вышки (м) | Размеры площадки (м) | Количество промежуточных секций | Цена | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
ПЕРЕДВИЖНАЯ СБОРНО-РАЗБОРНАЯ ВЫШКА (С ПОЛИМЕРНЫМ ПОКРЫТИЕМ) ПСРВ-7,5 | |||||||||||||||||||
| Вышка-тура ПСРВ-7.5 / 2.7 | 2.7 | 1.7 x 0.7 | 1 | 18,100.00 ₽ | Количество | ||||||||||||||
| Вышка-тура ПСРВ-7.5 / 3.9 | 3.9 | 1.7 x 0.7 | 2 | 19,700.00 ₽ | Количество | ||||||||||||||
| Вышка-тура ПСРВ-7.5 / 5.1 | 5.1 | 1.7 x 0.7 | 3 | 25,950.00 ₽ | Количество | ||||||||||||||
Вышка-тура ПСРВ-7. | 6.3 | 1.7 x 0.7 | 4 | 28,950.00 ₽ | Количество | ||||||||||||||
| Вышка-тура ПСРВ-7.5 / 7.5 | 7.5 | 1.7 x 0.7 | 5 | 32,600.00 ₽ | Количество | ||||||||||||||
| Базовый блок для вышки туры ПСРВ-7.5 | 13,800.00 ₽ | Количество | |||||||||||||||||
| Промежуточная секция для вышки туры ПСРВ-7.5 | 3,500.00 ₽ | Количество | |||||||||||||||||
ПЕРЕДВИЖНАЯ СБОРНО-РАЗБОРНАЯ ВЫШКА (С ПОЛИМЕРНЫМ ПОКРЫТИЕМ) ПСРВ-21 | |||||||||||||||||||
| Вышка-тура ПСРВ-21 / 2.7 | 2.7 | 2.00 x 1.20 | 1 | 21,990.00 ₽ | Количество | ||||||||||||||
| Вышка-тура ПСРВ-21 / 3.9 | 3.9 | 2.00 x 1.20 | 2 | 25,450. 00 ₽ 00 ₽ | Количество | ||||||||||||||
| Вышка-тура ПСРВ-21 / 5.1 | 5.1 | 2.00 x 1.20 | 3 | 32,100.00 ₽ | Количество | ||||||||||||||
| Вышка-тура ПСРВ-21 / 6.3 | 6.3 | 2.00 x 1.20 | 4 | 35,950.00 ₽ | Количество | ||||||||||||||
| Вышка-тура ПСРВ-21 / 7.5 | 7.5 | 2.00 x 1.20 | 5 | 39,850.00 ₽ | Количество | ||||||||||||||
| Вышка-тура ПСРВ-21 / 8.7 | 8.7 | 2.00 x 1.20 | 6 | 44,600.00 ₽ | Количество | ||||||||||||||
| Вышка-тура ПСРВ-21 / 9.9 | 9.9 | 2.00 x 1.20 | 7 | 48,000.00 ₽ | Количество | ||||||||||||||
| Вышка-тура ПСРВ-21 / 11.1 | 11.1 | 2.00 x 1.20 | 8 | 52,720.00 ₽ | Количество | ||||||||||||||
Вышка-тура ПСРВ-21 / 12. 3 3 | 12.3 | 2.00 x 1.20 | 9 | 56,870.00 ₽ | Количество | ||||||||||||||
| Вышка-тура ПСРВ-21 / 13.5 | 13.5 | 2.00 x 1.20 | 10 | 60,950.00 ₽ | Количество | ||||||||||||||
| Вышка-тура ПСРВ-21 / 14.7 | 14.7 | 2.00 x 1.20 | 11 | 64,900.00 ₽ | Количество | ||||||||||||||
| Вышка-тура ПСРВ-21 / 15.9 | 15.9 | 2.00 x 1.20 | 12 | 68,350.00 ₽ | Количество | ||||||||||||||
| Вышка-тура ПСРВ-21 / 17.1 | 17.1 | 2.00 x 1.20 | 13 | 73,000.00 ₽ | Количество | ||||||||||||||
| Вышка-тура ПСРВ-21 / 18.3 | 18.3 | 2.00 x 1.20 | 14 | 76,900.00 ₽ | Количество | ||||||||||||||
Вышка-тура ПСРВ-21 / 19. 5 5 | 19.5 | 2.00 x 1.20 | 15 | 79,950.00 ₽ | Количество | ||||||||||||||
| Вышка-тура ПСРВ-21 / 20.7 | 20.7 | 2.00 x 1.20 | 16 | 84,830.00 ₽ | Количество | ||||||||||||||
| Базовый блок для вышки туры ПСРВ-21 | 18,240.00 ₽ | Количество | |||||||||||||||||
| Промежуточная секция для вышки туры ПСРВ-21 | 4,200.00 ₽ | Количество | |||||||||||||||||
ПЕРЕДВИЖНАЯ СБОРНО-РАЗБОРНАЯ ВЫШКА (С ПОЛИМЕРНЫМ ПОКРЫТИЕМ) ПСРВ-22 | |||||||||||||||||||
| Вышка-тура ПСРВ-22 / 2.7 | 2.7 | 2.00 x 2.00 | 1 | 27,500.00 ₽ | Количество | ||||||||||||||
| Вышка-тура ПСРВ-22 / 3.9 | 3.9 | 2.00 x 2.00 | 2 | 29,950. 00 ₽ 00 ₽ | Количество | ||||||||||||||
| Вышка-тура ПСРВ-22 / 5.1 | 5.1 | 2.00 x 2.00 | 3 | 37,990.00 ₽ | Количество | ||||||||||||||
| Вышка-тура ПСРВ-22 / 6.3 | 6.3 | 2.00 x 2.00 | 4 | 41,980.00 ₽ | Количество | ||||||||||||||
| Вышка-тура ПСРВ-22 / 7.5 | 7.5 | 2.00 x 2.00 | 5 | 46,700.00 ₽ | Количество | ||||||||||||||
| Вышка-тура ПСРВ-22 / 8.7 | 8.7 | 2.00 x 2.00 | 6 | 50,880.00 ₽ | Количество | ||||||||||||||
| Вышка-тура ПСРВ-22 / 9.9 | 9.9 | 2.00 x 2.00 | 7 | 54,800.00 ₽ | Количество | ||||||||||||||
| Вышка-тура ПСРВ-22 / 11.1 | 11.1 | 2.00 x 2.00 | 8 | 59,900.00 ₽ | Количество | ||||||||||||||
Вышка-тура ПСРВ-22 / 12. 3 3 | 12.3 | 2.00 x 2.00 | 9 | 64,650.00 ₽ | Количество | ||||||||||||||
| Вышка-тура ПСРВ-22 / 13.5 | 13.5 | 2.00 x 2.00 | 10 | 68,100.00 ₽ | Количество | ||||||||||||||
| Вышка-тура ПСРВ-22 / 14.7 | 14.7 | 2.00 x 2.00 | 11 | 73,300.00 ₽ | Количество | ||||||||||||||
| Вышка-тура ПСРВ-22 / 15.9 | 15.9 | 2.00 x 2.00 | 12 | 78,400.00 ₽ | Количество | ||||||||||||||
| Вышка-тура ПСРВ-22 / 17.1 | 17.1 | 2.00 x 2.00 | 13 | 83,300.00 ₽ | Количество | ||||||||||||||
| Вышка-тура ПСРВ-22 / 18.3 | 18.3 | 2.00 x 2.00 | 14 | 92,450.00 ₽ | Количество | ||||||||||||||
Вышка-тура ПСРВ-22 / 19. 5 5 | 19.5 | 2.00 x 2.00 | 15 | 94,700.00 ₽ | Количество | ||||||||||||||
| Вышка-тура ПСРВ-22 / 20.7 | 20.7 | 2.00 x 2.00 | 16 | 96,200.00 ₽ | Количество | ||||||||||||||
| Вышка-тура ПСРВ-22 / 21.9 | 21.9 | 2.00 x 2.00 | 17 | 102,000.00 ₽ | Количество | ||||||||||||||
| Базовый блок для вышки туры ПСРВ-22 | 21,600.00 ₽ | Количество | |||||||||||||||||
| Промежуточная секция для вышки туры ПСРВ-22 | 4,800.00 ₽ | Количество | |||||||||||||||||
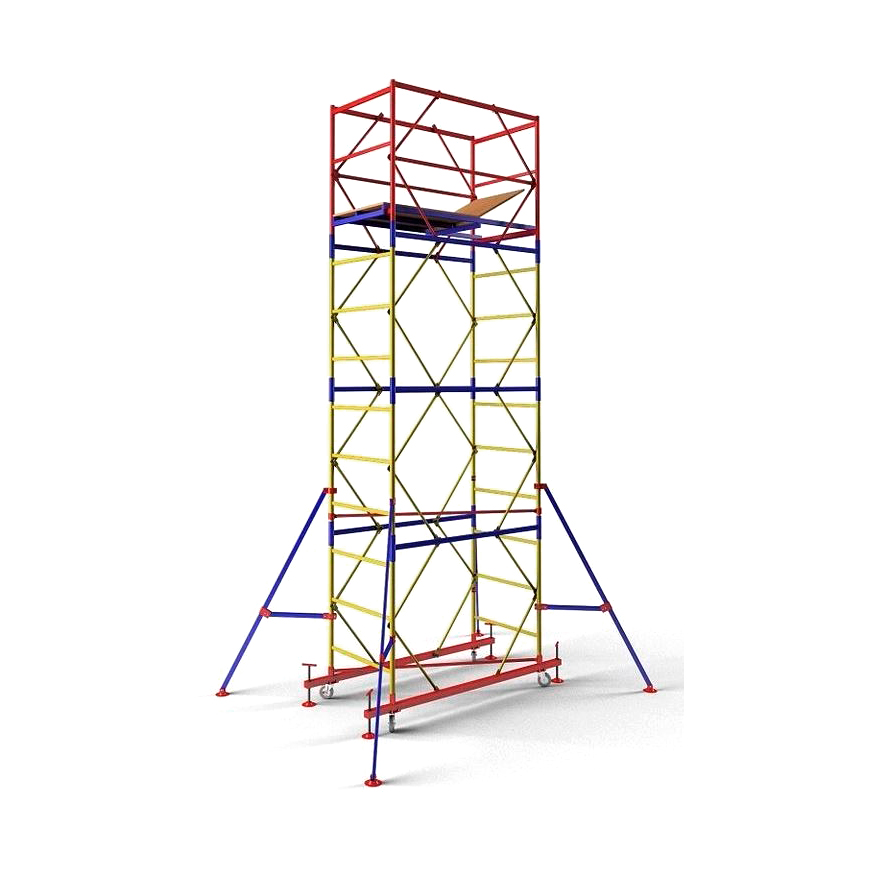
Вышка-тура ПСРВ-7,5
Вышка передвижная сборно-разборная ПСРВ-7,5 (с полимерным покрытием).
Вышка-тура ПСРВ-7,5 с размером рабочей площадки 1,67х0,8 метра идеальна для проведения электромонтажных, строительных, эксплуатационных и ремонтных работ на высоте от 3 до 21 метра. Работы можно вести как внутри помещений, так и на открытых площадках.
Конструкция передвижной сборно-разборной вышки надежна, основные элементы её изготовлены из стальной трубы диаметром 42 миллиметра, не основные элементы – из трубы 25мм., рабочая площадка оборудована настилами из влагостойкой фанеры (2 настила – с люком и без люка). Покрытие всех металлических элементов вышки – полимерно-порошковое, препятствует преждевременной коррозии, увеличивает срок службы. Для обеспечения устойчивости, тура снабжена уравнивающими опорами и раскосами, для передвижения по опорной поверхности у вышки предусмотрены колеса. Для обеспечения безопасных условий труда рабочая площадка имеет защитное ограждение.
Вышка-тура ПСРВ легко собирается из стандартных секций высотой 1,2 метра, раскрепленных горизонтальными и диагональными связями. Крепление элементов – флажковое. Быстрый монтаж и демонтаж без использования инструментов позволяет монтажникам собирать их в течение 15 минут. Нагрузка на рабочий настил вышки – 250кг, статическая нагрузка на перила ограждения – 70кг.
Крепление элементов – флажковое. Быстрый монтаж и демонтаж без использования инструментов позволяет монтажникам собирать их в течение 15 минут. Нагрузка на рабочий настил вышки – 250кг, статическая нагрузка на перила ограждения – 70кг.
Вышка передвижная сборно-разборная ПСРВ-7,5 имеет следующие технические параметры:
|
Модель вышки |
Количество промежуточных секций, шт |
Полная высота |
Максимальная высота установки настила, м. |
Размеры рабочей площадки, м |
Масса, кг |
|
Вышка ПСРВ-7,5-2,69 |
1 |
2,69 |
1,69 |
1,67х0,8 |
135,9 |
|
Вышка ПСРВ-7,5-3,89 |
2 |
3,89 |
2,89 |
1,67х0,8 |
160,9 |
|
Вышка ПСРВ-7,5-5,09 |
3 |
5,09 |
4,09 |
1,67х0,8 |
185,9 |
|
Вышка ПСРВ-7,5-6,29 |
4 |
6,29 |
5,29 |
1,67х0,8 |
210,9 |
|
Вышка ПСРВ-7,5-7,49 |
5 |
7,49 |
6,49 |
1,67х0,8 |
235,9 |
|
Промежуточная секция |
1,2 |
|
|
|
25,0 |
Дополнительную информацию по модели Вышка передвижная сборно-разборная ПСРВ-7,5: описание, цена, технические параметры, наличие на складе необходимо уточнять у поставщика в г. Ижевск — компании «ИжПодъем», по телефону +7 (3412) 55-20-25 или по электронной почте
Этот адрес электронной почты защищен от спам-ботов. У вас должен быть включен JavaScript для просмотра.
Ижевск — компании «ИжПодъем», по телефону +7 (3412) 55-20-25 или по электронной почте
Этот адрес электронной почты защищен от спам-ботов. У вас должен быть включен JavaScript для просмотра.
- Вышка передвижная сборно-разборная ПСРВ-21
- Вышка передвижная сборно-разборная ПСРВ-22
Неперсистентный потивирус, переносимый тлей, по-разному изменяет биологию переносчика и непереносчика посредством манипуляций с качеством растения-хозяина
1. Eigenbrode SD, Ding H, Shiel P, Berger PH. Летучие вещества растений картофеля, инфицированных вирусом скручивания листьев картофеля , привлекают и блокируют переносчик вируса Myzus persicae (Homoptera: Aphididae) Proc. Р. Соц. Лонд. [Биол.]. 2002; 269: 455–460. doi: 10.1098/rspb.2001.1909. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
2. Mauck KE, Bosque-Pérez NA, Eigenbrode SD, Moraes CM, Mescher MC. Механизмы передачи определяют влияние патогенов на взаимодействия хозяина и переносчика: данные вирусов растений. Функц. Экол. 2012;26:1162–1175. doi: 10.1111/j.1365-2435.2012.02026.x. [CrossRef] [Google Scholar]
Механизмы передачи определяют влияние патогенов на взаимодействия хозяина и переносчика: данные вирусов растений. Функц. Экол. 2012;26:1162–1175. doi: 10.1111/j.1365-2435.2012.02026.x. [CrossRef] [Google Scholar]
3. Bosque-Pérez NA, Eigenbrode SD. Влияние вызванных вирусом изменений в растениях на переносчиков тлей: выводы из патосистем лютеовирусов. Вирус рез. 2011; 159: 201–205. doi: 10.1016/j.virusres.2011.04.020. [PubMed] [CrossRef] [Академия Google]
4. Ingwell, L.L., Eigenbrode, S.D. & Bosque-Pérez, N.A. Вирусы растений изменяют поведение насекомых, способствуя их распространению. Науч. Реп , 10.1038/srep00578 (2012). [Бесплатная статья PMC] [PubMed]
5. Eigenbrode SD, Bosque-Pérez NA, Davis TS. Переносимые насекомыми патогены растений и их переносчики: экология, эволюция и сложные взаимодействия. Анну. Преподобный Энтомол. 2018;63:169–191. doi: 10.1146/annurev-ento-020117-043119. [PubMed] [CrossRef] [Google Scholar]
6. Карр, Дж. П. и др. . Вирусные манипуляции с реакцией растений на стресс и взаимодействие хозяина с насекомыми в Advances in Virus Research Vol. 102 , 177–197, 10.1016/bs.aivir.2018.06.004, (Elsevier, 2018). [PubMed]
. Вирусные манипуляции с реакцией растений на стресс и взаимодействие хозяина с насекомыми в Advances in Virus Research Vol. 102 , 177–197, 10.1016/bs.aivir.2018.06.004, (Elsevier, 2018). [PubMed]
7. Mauck, K.E., Chesnais, Q. & Shapiro, L.R. Эволюционные детерминанты манипулирования хозяином и вектором вирусами растений. В Advances in Virus Research Vol. 101, 189–250, 10.1016/bs.aivir.2018.02.007 (Elsevier, 2018). [PubMed]
8. Mauck KE, De Moraes CM, Mescher MC. Биохимические и физиологические механизмы, лежащие в основе эффектов Вирус мозаики огурца на признаки растения-хозяина, которые опосредуют передачу тлей-переносчиками. Окружающая среда растительной клетки. 2014; 37:1427–1439. doi: 10.1111/pce.12249. [PubMed] [CrossRef] [Google Scholar]
9. Blua MJ, Perring TM, Madore MA. Вызванные вирусом растений изменения в развитии популяции тли и временные колебания питательных веществ растений. Дж. Хим. Экол. 1994; 20: 691–707. doi: 10.1007/BF02059607. [PubMed] [CrossRef] [Google Scholar]
1994; 20: 691–707. doi: 10.1007/BF02059607. [PubMed] [CrossRef] [Google Scholar]
10. Бокель С., Джорданенго П., Амелин А. Дивергентное воздействие картофеля, инфицированного PVY, на тлю. Евро. Дж. Плант Патол. 2011;129: 507–510. doi: 10.1007/s10658-010-9732-8. [CrossRef] [Google Scholar]
11. Salvaudon L, De Moraes CM, Mescher MC. Исходы коинфекции двумя потивирусами: значение для эволюции манипулятивных стратегий. проц. Р. Соц. Б. 2013; 280:20122959. doi: 10.1098/rspb.2012.2959. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
12. Casteel CL, et al. Белок NIa-Pro вируса мозаики турнепса улучшает рост и размножение тли-переносчика Myzus persicae (зеленая персиковая тля) Plant J. 2014;77:653–663. doi: 10.1111/tpj.12417. [PubMed] [CrossRef] [Google Scholar]
13. Casteel, C. et al . Нарушение реакции на этилен вирусом мозаики турнепса опосредует подавление защиты растений от тли-переносчика Myzus persicae . Завод Физиол , 10.1104/с.15.00332 (2015). [Статья бесплатно PMC] [PubMed]
Завод Физиол , 10.1104/с.15.00332 (2015). [Статья бесплатно PMC] [PubMed]
14. Бак, А., Чунг, А. Л., Ян, К., Уитэм, С. А. и Кастил, С. Л. Вирусная протеаза перемещается в присутствии вектора, чтобы повысить эффективность вектора. Нац. Сообщение , 10.1038/ncomms14493 (2017). [Бесплатная статья PMC] [PubMed]
15. Mauck KE, De Moraes CM, Mescher MC. Обманчивые химические сигналы, индуцируемые растительным вирусом, привлекают насекомых-переносчиков к низшим хозяевам. проц. Натл. акад. науч. США. 2010;107:3600–3605. doi: 10.1073/pnas.0907191107. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
16. Zhang T, et al. Begomovirus – мутуализм белокрылки достигается за счет подавления защитных сил растений фактором патогенности вируса. Мол. Экол. 2012;21:1294–1304. doi: 10.1111/j.1365-294X.2012.05457.x. [PubMed] [CrossRef] [Google Scholar]
17. Шринивасан Р., Альварес Дж. М., Эйгенброде С. Д., Боске-Перес Н. А. Влияние паслена мохнатого Solanum sarrachoides (Sendtner) и вируса скручивания листьев картофеля (Luteoviridae: Polerovirus) на предпочтение хозяина Myzus persicae (Sulzer) (Homoptera: Aphididae) Environ. Энтомол. 2006; 35: 546–553. doi: 10.1603/0046-225X-35.2.546. [CrossRef] [Google Scholar]
Энтомол. 2006; 35: 546–553. doi: 10.1603/0046-225X-35.2.546. [CrossRef] [Google Scholar]
18. Пикетт Дж. А., Уодхамс Л. Дж., Вудкок С. М., Харди Дж. Химическая экология тлей. Анну. Преподобный Энтомол. 1992;37:67–90. doi: 10.1146/annurev.en.37.010192.000435. [CrossRef] [Google Scholar]
19. Хукс CRR, Феререс А. Защита сельскохозяйственных культур от вирусов, переносимых непостоянной тлей: обзор использования барьерных растений в качестве инструмента управления. Вирус рез. 2006; 120:1–16. doi: 10.1016/j.virusres.2006.02.006. [PubMed] [CrossRef] [Google Scholar]
20. Деринг, Т. Ф. и Читтка, Л. Визуальная экология тли — критический обзор роли цветов в поиске хозяина. Взаимодействие членистоногих и растений . 10.1007/s11829-006-9000-1 (2007).
21. Хименес-Мартинес Э.С. и др. Летучие сигналы влияют на реакцию Rhopalosiphum padi (Homoptera: Aphididae) на вируса желтой карликовости ячменя – инфицированную трансгенную и нетрансформированную пшеницу. Окружающая среда. Энтомол. 2004; 33: 1207–1216. doi: 10.1603/0046-225X-33.5.1207. [CrossRef] [Google Scholar]
Окружающая среда. Энтомол. 2004; 33: 1207–1216. doi: 10.1603/0046-225X-33.5.1207. [CrossRef] [Google Scholar]
22. Феререс А., Кампмайер Г., Ирвин М. Привлекательность и предпочтение тли для зараженных растений сои и перца. Потивирусы. Анна. Энтомол. соц. Являюсь. 1999;92:542–548. doi: 10.1093/aesa/92.4.542. [CrossRef] [Google Scholar]
23. Дональдсон Дж. Р., Граттон С. Антагонистические эффекты соевых вирусов на продуктивность соевой тли. Окружающая среда. Энтомол. 2007; 36: 918–925. doi: 10.1093/ee/36.4.918. [PubMed] [CrossRef] [Google Scholar]
24. Марккула М., Лаурема С. Изменения концентрации свободных аминокислот в растениях, индуцированные вирусными заболеваниями и репродукцией тли. Annales Agriculturae Fenniae. 1964; 3: 265–271. [Академия Google]
25. Аджайи О. Влияние вируса желтой карликовости ячменя на аминокислотный состав яровой пшеницы. Анна. заявл. биол. 1986; 108: 145–149. doi: 10.1111/j.1744-7348.1986.tb01975.x. [CrossRef] [Google Scholar]
26. Fereres A, Shukle R, Araya J, Foster J. Зондирование и кормление Sitobion avenae (F.) (Horn., Aphididae) на трех сортах пшеницы, инфицированных Barley. вирус желтой карликовости . Дж. Заявл. Энтомол. 1990; 109: 29–36. дои: 10.1111/j.1439-0418.1990.tb00015.x. [CrossRef] [Google Scholar]
Fereres A, Shukle R, Araya J, Foster J. Зондирование и кормление Sitobion avenae (F.) (Horn., Aphididae) на трех сортах пшеницы, инфицированных Barley. вирус желтой карликовости . Дж. Заявл. Энтомол. 1990; 109: 29–36. дои: 10.1111/j.1439-0418.1990.tb00015.x. [CrossRef] [Google Scholar]
27. Аджайи О., Дьюар А. Влияние вируса желтой карликовости ячменя на полевые популяции злаковых тлей, Sitobion avenae и Metopolophium dirhodum . Анна. заявл. биол. 1983; 103:1–11. doi: 10.1111/j.1744-7348.1983.tb02734.x. [CrossRef] [Google Scholar]
28. Castle S, Berger P. Темпы роста и прироста Myzus persicae на зараженном вирусом картофеле в зависимости от типа взаимоотношений вирус-переносчик. Энтомол. Эксп. заявл. 1993;69:51–60. doi: 10.1111/j.1570-7458.1993.tb01727.x. [CrossRef] [Google Scholar]
29. Eckel W, Rand V, Lampert E. Относительная привлекательность зараженных вирусом травления табака и здоровых растений табака, подвергнутых дымовой сушке, для тли (Homoptera: Aphididae) J. Econ. Энтомол. 1996; 89: 1017–1027. doi: 10.1093/jee/89.4.1017. [CrossRef] [Google Scholar]
Econ. Энтомол. 1996; 89: 1017–1027. doi: 10.1093/jee/89.4.1017. [CrossRef] [Google Scholar]
30. Blua M, Perring T. Производство и увеличение популяции крылатых тлей-переносчиков на зараженных вирусом растениях-хозяевах. Экология. 1992; 92: 65–70. doi: 10.1007/BF00317263. [PubMed] [CrossRef] [Академия Google]
31. Yeh SD, Gonsalves D, Provvidenti R. Сравнительные исследования диапазона хозяев и серологии вируса кольцевой пятнистости папайи и вируса мозаики арбуза 1 . Фитопатология. 1984; 74: 1081–1085. doi: 10.1094/Phyto-74-1081. [CrossRef] [Google Scholar]
32. Kalleshwaraswamy C, Kumar NK. Эффективность передачи вируса кольцевой пятнистости папайи тремя видами тлей. Фитопатология. 2008; 98: 541–546. doi: 10.1094/PHYTO-98-5-0541. [PubMed] [CrossRef] [Академия Google]
33. Кастл С., Моури Т., Бергер П. Дифференциальное заселение Myzus persicae (Homoptera: Aphididae) на различных зараженных вирусом растениях-хозяевах. Анна. Энтомол. соц. Являюсь. 1998; 91: 661–667. doi: 10.1093/aesa/91.5.661. [CrossRef] [Google Scholar]
Анна. Энтомол. соц. Являюсь. 1998; 91: 661–667. doi: 10.1093/aesa/91.5.661. [CrossRef] [Google Scholar]
34. Шринивасан Р., Альварес Дж. М. Влияние смешанных вирусных инфекций ( Potatovirus Y–Potato leafletvirus ) на биологию и предпочтение переносчиков Myzus persicae и Macrosiphum euphorbiae (Hemiptera: Aphididae) J. Econ. Энтомол. 2007; 100: 646–655. дои: 10.1603/0022-0493(2007)100[646:EOMVIP]2.0.CO;2. [PubMed] [CrossRef] [Google Scholar]
35. Ziebell, H. et al . Вирус мозаики огурца и его РНК-супрессор 2b модифицируют взаимодействие растений и тли в табаке. Науч. Реп , 10.1038/srep00187 (2011). [Бесплатная статья PMC] [PubMed]
36. Jiménez J, et al. Голодание изменяет поведение тли при зондировании, но не всегда увеличивает скорость передачи нециркулирующих вирусов. Дж. Генерал Вирол. 2017;98:3111–3121. дои: 10.1099/jgv.0.000971. [PubMed] [CrossRef] [Google Scholar]
37. Hewer A, Will T, van Bel AJ. Растительные сигналы для навигации тли в сосудистых тканях. Дж. Эксп. биол. 2010; 213:4030–4042. doi: 10.1242/jeb.046326. [PubMed] [CrossRef] [Google Scholar]
Растительные сигналы для навигации тли в сосудистых тканях. Дж. Эксп. биол. 2010; 213:4030–4042. doi: 10.1242/jeb.046326. [PubMed] [CrossRef] [Google Scholar]
38. Пауэлл Г., Тош Ч.Р., Харди Дж. Выбор растений-хозяев тлей: поведенческие, эволюционные и прикладные перспективы. Анну. Преподобный Энтомол. 2006; 51: 309–330. doi: 10.1146/annurev.ento.51.110104.151107. [PubMed] [CrossRef] [Академия Google]
39. Оклер Ж.Л. Кормление и питание тли. Анну. Преподобный Энтомол. 1963; 8: 439–490. doi: 10.1146/annurev.en.08.010163.002255. [CrossRef] [Google Scholar]
40. Миттлер Т. Влияние концентрации аминокислот и сахаров на поглощение пищи тлей Myzus persicae . Энтомол. Эксп. заявл. 1967; 10: 39–51. doi: 10.1111/j.1570-7458.1967.tb00043.x. [CrossRef] [Google Scholar]
41. Mittler T, Dadd R, Daniels S. Использование различных сахаров тлей Myzus persicae . J. Физиология насекомых. 1970; 16: 1873–1890. doi: 10.1016/0022-1910(70)-9. [CrossRef] [Google Scholar]
42. Эмден Х. В. и Башфорд, Массачусетс. Эффективность Brevicoryne brassicae и Myzus persicae по отношению к возрасту растений и содержанию аминокислот в листьях. Энтомол. Эксп. заявл. 1971; 14: 349–360. doi: 10.1111/j.1570-7458.1971.tb00172.x. [CrossRef] [Google Scholar]
Эмден Х. В. и Башфорд, Массачусетс. Эффективность Brevicoryne brassicae и Myzus persicae по отношению к возрасту растений и содержанию аминокислот в листьях. Энтомол. Эксп. заявл. 1971; 14: 349–360. doi: 10.1111/j.1570-7458.1971.tb00172.x. [CrossRef] [Google Scholar]
43. Febvay G, et al. Устойчивость различных сортов люцерны к гороховой тле Acyrthosiphon pisum : влияние состава флоэмы на плодовитость тлей. Энтомол. Эксп. заявл. 1988; 48: 127–134. doi: 10.1111/j.1570-7458.1988.tb01155.x. [CrossRef] [Google Scholar]
44. Sandström J, Pettersson J. Аминокислотный состав флоэмного сока и связь с внутривидовой изменчивостью у гороховой тли ( Acyrthosiphon pisum ). J. Физиология насекомых. 1994; 40:947–955. doi: 10.1016/0022-1910(94)-3. [CrossRef] [Google Scholar]
45. Карли А., Дуглас А., Паркер В. Аминокислотный состав и пищевая ценность сока флоэмы листьев картофеля для борьбы с тлей. Дж. Эксп. биол. 2002;205:3009–3018.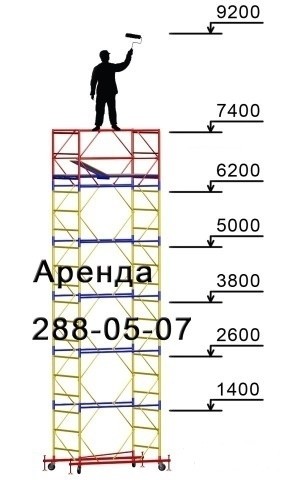 [PubMed] [Google Scholar]
[PubMed] [Google Scholar]
46. Дуглас А.Е. Пищевая физиология тли. Доп. Физиол насекомых. 2003; 31: 73–140. doi: 10.1016/S0065-2806(03)31002-1. [CrossRef] [Google Scholar]
47. Maltais J, Auclair J. Композиция свободных аминокислот и амидов сока листьев гороха, гемолимфы гороховой тли и пади после выращивания тли на отдельных листьях гороха, обработанных аминосоединениями. J. Физиология насекомых. 1962; 8: 391–399. doi: 10.1016/0022-1910(62)
-2. [Перекрестная ссылка] [Академия Google]48. Ретнакаран А., Бек С.Д. Потребность в аминокислотах и метаболизм сернистых аминокислот у гороховой тли, Acyrthosiphon pisum (Harris) Comp. Биохим. Физиол. 1968; 24: 611–619. doi: 10.1016/0010-406X(68)91013-X. [PubMed] [CrossRef] [Google Scholar]
49. Сандстрем, Дж. и Моран, Н. Насколько несбалансирован питательный сок флоэмы для тли? В Материалы 10-го Международного симпозиума по взаимоотношениям насекомых и растений. Series Entomologica , Vol 56, 203-210 (Springer, 1999).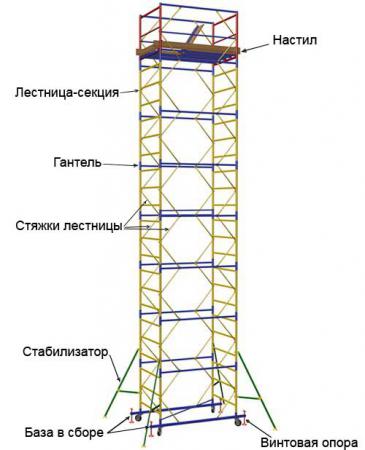
50. Febvay G, Rahbé Y, Rynkiewicz M, Guillaud J, Bonnot G. Судьба диетической сахарозы и неосинтез аминокислот у гороховой тли, Acyrthosiphon pisum , выращенных на разных диетах. Дж. Эксп. биол. 1999; 202: 2639–2652. [PubMed] [Google Scholar]
51. Эшфорд Д., Смит В., Дуглас А. Жизнь на диете с высоким содержанием сахара: судьба сахарозы, проглоченной насекомым, питающимся флоэмой, гороховой тлей Acyrthosiphon pisum . J. Физиология насекомых. 2000;46:335–341. дои: 10.1016/S0022-1910(99)00186-9. [PubMed] [CrossRef] [Google Scholar]
52. Fiehn O. Метаболические сети Cucurbita maxima флоэмы. Фитохимия. 2003; 62: 875–886. doi: 10.1016/S0031-9422(02)00715-X. [PubMed] [CrossRef] [Google Scholar]
53. Abisgold JD, Simpson SJ, Douglas AE. Регуляция питательных веществ у гороховой тли Acyrthosiphon pisum: применение новой геометрической схемы к потреблению сахара и аминокислот. Физиол. Энтомол. 1994; 19: 95–102. дои: 10. 1111/j.1365-3032.1994.tb01081.х. [CrossRef] [Google Scholar]
1111/j.1365-3032.1994.tb01081.х. [CrossRef] [Google Scholar]
54. Ван Р., Габриал С. Влияние поведения тли на эффективность передачи вируса мозаики сои тлей, колонизирующей сою, Aphis glycines . Заболевание растений. 2002; 86: 1260–1264. doi: 10.1094/PDIS.2002.86.11.1260. [CrossRef] [Google Scholar]
55. Ng JC, Falk BW. Взаимодействия вирус-вектор, опосредующие непостоянную и полупостоянную передачу вирусов растений. Анну. Преподобный Фитопат. 2006; 44: 183–212. doi: 10.1146/annurev.phyto.44.070505.143325. [PubMed] [CrossRef] [Академия Google]
56. Саад, К. А., Мохамад Рофф, М., Халлетт, Р. Х. и АБд-Гани, И. Б. Воздействие вируса мозаики огурца , инфицированного растениями перца чили, на растения, не являющиеся переносчиками Bemisia tabaci (Hemiptera: Aleyrodidae). Наука о насекомых . (2017). [PubMed]
57. Belliure B, Sabelis MW, Janssen A. Вектор и вирус вызывают реакции растений, которые приносят пользу травоядным, не являющимся переносчиками. Базовое приложение Экол. 2010; 11: 162–169. doi: 10.1016/j.baae.2009.09.004. [CrossRef] [Google Scholar]
Базовое приложение Экол. 2010; 11: 162–169. doi: 10.1016/j.baae.2009.09.004. [CrossRef] [Google Scholar]
58. Анхелес-Лопес Ю.И., Ривера-Бустаманте Р., Хейл М. Фатальное привлечение непереносчиков ухудшает приспособленность манипулирующего растительного вируса. Дж. Экол. 2018;106:391–400. doi: 10.1111/1365-2745.12838. [CrossRef] [Google Scholar]
59. Blackmer J, Byrne D. Изменения аминокислот в Cucumis melo в связи с особенностями жизненного цикла и склонностью к бегству Bemisia tabaci . Энтомол. Эксп. заявл. 1999; 93: 29–40. doi: 10.1046/j.1570-7458.1999.00559.x. [CrossRef] [Google Scholar]
60. Бирн Д.Н., Миллер В.Б. Углеводный и аминокислотный состав флоэмного сока и пади производства Bemisia tabaci . J. Физиология насекомых. 1990;36:433–439. doi: 10.1016/0022-1910(90)-J. [CrossRef] [Google Scholar]
61. Картер, Н. и Харрингтон, Р. Достижения в исследованиях переносчиков болезней . 19–51 (Спрингер, 1991).
62. Легарреа С., Барман А., Маршан В., Диффи С., Шринивасан Р. Временные эффекты инфекции Begomovirus и устойчивости растений-хозяев на предпочтение и развитие насекомых-переносчиков, Bemisia tabaci и последствия для эпидемий . ПЛОС Один. 2015;10:e0142114. doi: 10.1371/journal.pone.0142114. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
63. Wyatt, I.J. & White, P.F. Простая оценка внутренних темпов роста тлей и тетранихидных клещей. J. Appl. Экол . 757–766 (1977).
64. Santander J, et al. Механизмы внутренней устойчивости к антимикробным пептидам Edwardsiella ictaluri и ее влияние на воспаление и вирулентность кишечника рыб. Микробиология. 2013; 159:1471–1486. doi: 10.1099/микрофон 0.066639-0. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
65. Bolker BM, et al. Обобщенные линейные смешанные модели: практическое руководство по экологии и эволюции. Тенденции Экол. Эвол. 2009 г.;24:127–135. doi: 10.1016/j.tree.2008.10.008. [PubMed] [CrossRef] [Google Scholar]
doi: 10.1016/j.tree.2008.10.008. [PubMed] [CrossRef] [Google Scholar]
Экологичность здания
Экологичность здания
Мы спроектировали наше здание с учетом экологической устойчивости. Первоначальная цель состояла в том, чтобы получить бронзовый сертификат LEED. К сожалению, стоимость сертификации вышла за рамки нашего бюджета, но мы все равно продвинулись вперед. Наше здание подчеркивает две ключевые темы устойчивого развития – сокращение и повторное использование. Мы выделили все зоны в нашем здании, где были реализованы эти две темы.
Пивоварня
1. Резервуар для холодных напитков. Дядя владельца и один из наших фермеров подарили нам свой старый резервуар для хранения молока, в котором ферма хранила молоко, когда работала как молочная ферма. Этот танк в семье с 1979 года и прослужил в хозяйстве 27 лет. Мы гордимся тем, что используем этот резервуар на нашем пивоваренном заводе.
2. Охлаждающая вода. Резервуар для наливных молочных продуктов позволяет пивоварне экономить воду, потому что мы сохраняем и повторно используем воду, которая использовалась для охлаждения нашего пива в процессе пивоварения.
3. Резервуар для холодного спирта. Предварительно охладив нашу охлаждающую воду перед днем заваривания, мы можем медленно охлаждать воду в течение ночи во время непикового использования энергии. Это помогает снизить пиковую нагрузку на нашу электростанцию, что означает более эффективное производство электроэнергии.
4. Дробь. Мы сохраняем дробину, оставшуюся после варки пива, и отправляем ее нашим фермерам. Зерно по-прежнему содержит ценные белки и питательные вещества, которые служат полезным источником пищи для животных. Когда фермеры собирают зерно вскоре после варки, оно еще теплое, и животные получают горячую еду (которую они любят так же сильно, как мы с вами)!
5. Приготовление пива встык. Когда это возможно, мы варим несколько партий пива последовательно, что позволяет нам экономить энергию. Варка вплотную друг к другу позволяет нам сохранить остаточную тепловую энергию от процесса кипячения и охлаждения в партии 1 и использовать ее в партии 2, сокращая потребность в тепле во второй партии.
6. Нет транспорта — поскольку наше пиво производится на месте нам не нужно транспортировать его к конечному потребителю, и мы экономим топливо и энергию.
7. Без упаковки. Большая часть нашего пива подается прямо из больших резервуаров из нержавеющей стали в ваш бокал. Это означает, что нам не нужно потреблять материалы и энергию для упаковки нашего пива.
Кухня
8. Посудомоечная машина Energy Star – Наша посудомоечная машина – СОВРЕМЕННАЯ. Она потребляет значительно меньше энергии и воды, чем стандартная профессиональная посудомоечная машина. На мытье посуды может уйти 2/3 всей воды, используемой рестораном. Вот почему мы инвестировали в эффективную машину, которая использует вдвое меньше воды и энергии. Наша машина Hobart Advansys улавливает выходящий пар для предварительного нагрева поступающей воды, тем самым уменьшая количество горячей воды, необходимой машине.
9. Льдогенератор Energy Star — потребляет меньше энергии, чем стандартный льдогенератор.
10. Фритюрница EnerLogic — потребляет меньше энергии, чем стандартная фритюрница, и была оценена Калифорнийской энергетической комиссией как энергосберегающая.
11. Масло для фритюрницы. Масло для фритюрницы передается третьей стороне для переработки для других целей, в основном в качестве источника топлива.
12. Печь EnergyStar. Потребляет меньше энергии, чем стандартная духовка, и была признана Энергетической комиссией Калифорнии как энергосберегающая.
13. Клапан ополаскивания с низким расходом. В нашем помещении для мытья посуды мы используем распылительный клапан предварительного ополаскивания с низким расходом (менее 1,5 галлонов в минуту) (PRSV), чтобы очистить посуду от сильного мусора перед тем, как пропустить ее через нашу посудомоечную машину. Стандартные PRSV могут использовать 2-5 галлонов в минуту и потреблять в два раза больше воды, чем посудомоечная машина.
14. Система вытяжных колпаков. Наши вытяжные колпаки на кухне оснащены современной системой управления, которая активируется в зависимости от тепловой нагрузки. Это означает, что вентиляторы вообще не работают или работают медленнее, если кухня работает медленно. Это экономит электроэнергию и уменьшает количество необходимого кондиционированного подпиточного воздуха. Традиционные вытяжные системы работают непрерывно в течение дня, требуя, чтобы система HVAC тратила энергию на кондиционирование воздуха, который не нужно выпускать, а также мощность для работы вентиляторов.
Это означает, что вентиляторы вообще не работают или работают медленнее, если кухня работает медленно. Это экономит электроэнергию и уменьшает количество необходимого кондиционированного подпиточного воздуха. Традиционные вытяжные системы работают непрерывно в течение дня, требуя, чтобы система HVAC тратила энергию на кондиционирование воздуха, который не нужно выпускать, а также мощность для работы вентиляторов.
15. Холодильная стойка. У нас есть единая система охлаждения, которая обеспечивает все наши потребности в охлаждении. В большинстве ресторанов используются независимые блоки, где каждый блок имеет собственную независимую систему охлаждения. У нас много холодильников, если подумать о каждом холодильнике за барной стойкой, каждом холодильнике и холодильнике на кухне, льдогенераторе и каждом холодильнике или морозильной камере. С одним блоком, обеспечивающим охлаждение для всех наших потребностей, он может работать намного эффективнее, что снижает потребление энергии.
16. Местное мясо. Наше мясо поступает прямо с фермы к мяснику, что сокращает расстояние, которое оно преодолевает, и, следовательно, снижает потребление энергии, необходимой для транспортировки, например, ископаемого топлива.
Местное мясо. Наше мясо поступает прямо с фермы к мяснику, что сокращает расстояние, которое оно преодолевает, и, следовательно, снижает потребление энергии, необходимой для транспортировки, например, ископаемого топлива.
Здание
17. Восстановленный деревянный сарай. Когда владелец узнал о старом сарае, который собирались снести, он пошел (с помощью своей семьи) и удалил часть обшивки сарая, поэтому его можно было бы повторно использовать в Engrained для обшивки стен.
18. Восстановленное зернохранилище. Наше зернохранилище, стоящее за пределами нашей пивоварни, было получено с фермы семьи владельца. Они были так любезны, что привезли его для нас в Спрингфилд!
19. Застекленные окна. Остекление для этого проекта будет с низким коэффициентом излучения, сохраняющим тепло летом и зимой.
20. Толстая теплоизоляция стен. Наши стены хорошо изолированы (коэффициент R более 30 в большинстве мест), что сводит к минимуму потери тепла и охлаждения.
21. Маркизы на окнах. Над нашими окнами установлены навесы, предназначенные для защиты от света летом и пропускания света зимой. Летом угол наклона солнца к небу выше, а зимой угол наклона солнца к небу ниже. Это означает, что навес с правильными физическими размерами может создавать тень летом и пропускать теплый солнечный свет зимой.
22. ОВКВ. Мы установили высокоэффективные (рейтинг SEER 15,6) крышные блоки, управляемые усовершенствованной системой управления, использующей зональный контроль для эффективного регулирования здания.
23. Высокоэффективный водонагреватель. Эффективность нашего водонагревателя составляет 96 %, что позволяет максимально использовать природный газ, используемый для его топлива. Это лучший в отрасли показатель эффективности.
24. Двери вестибюля. Наличие «воздушного затвора» между двумя дверями предотвращает проникновение наружного воздуха при открытии двери. Когда снаружи есть единственная дверь, воздух быстро проникает в помещение, заставляя систему HVAC работать усерднее, чтобы кондиционировать воздух. Все наши наружные двери выполнены в стиле вестибюля, что обеспечивает экономию энергии по сравнению с традиционными дверями.
Все наши наружные двери выполнены в стиле вестибюля, что обеспечивает экономию энергии по сравнению с традиционными дверями.
25. Сушилки для рук в ванной. Наши сушилки для рук Xlerator экономят энергию, не требуют особого обслуживания и сокращают количество отходов. Они потребляют на 80% меньше энергии, чем традиционные сушилки для рук, и имеют меньший углеродный след по сравнению с бумажными полотенцами.
26. Фасеты автоматической раковины. Эти смесители предотвращают случайное оставление воды включенной.
27. Эффективные туалеты. В этих устройствах на один смыв расходуется меньше воды, чем в традиционных туалетах.
28. Светодиодные парковочные фонари. Наши парковочные фонари — это светодиоды, которые потребляют на 50 % меньше энергии, чем следующее лучшее решение — металлогалогенные лампы. Помимо экономии энергии, светодиодные фонари служат в 4+ раза дольше, прежде чем их нужно будет заменить.
29. Наружное светодиодное освещение. Во всех наших наружных светильниках, включая наши вывески, используется светодиодное освещение, которое потребляет наименьшее количество электроэнергии и служит дольше, чем любой другой источник света.