Аренда крана на гусеничном ходу МКГС-100 до 100 тонн в Санкт-Петербурге
Кран МКГС-100 — стреловой монтажный гусеничный кран, предназначенный для монтажа тяжеловесного и крупноблочного оборудования, строительных конструкций, а также реконструкции предприятий различных отраслей промышленности, включая металлургическую и нефтехимическую. Благодаря Г-образной компоновке башенно-стрелового оборудования очень удобен при большепролетном строительстве и с легкостью заменяет башенный кран, под установку которого требуется специальный фундамент. При работе на больших вылетах или через парапеты зданий в несколько раз дешевле подходящих автокранов.
Технические характеристики крана МКГС-100
- Максимальная грузоподъемность 100 тн
- Максимальная высота подъема 96 м
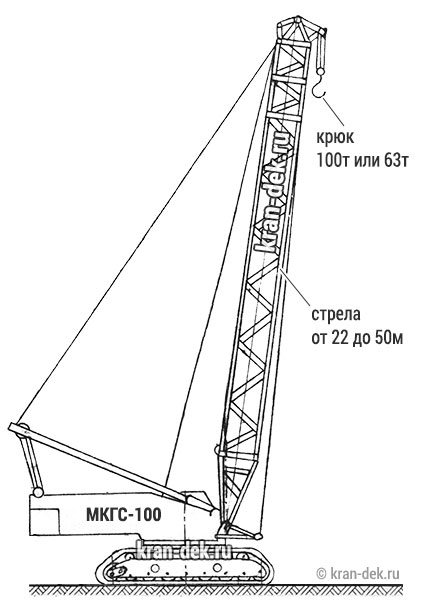
- Базовая длина стрелы 22 м
- Максимальная длина стрелы 57 м
- Максимальная длина маневрового гуся 40м
- Ширина ходового устройства 7000 мм
- Длина 9100 мм
- Высота 4250 мм
- Номинальная мощность двигателя 100 кВт
Конструкция крана МКГС-100 включает в себя ходовую и поворотную части, стреловое и башенно-стреловое оборудование, лебедку, крюковые обоймы, дизель-электрический агрегат, механизм вращения, монтажную стойку, генераторную группу и кабину опорно-поворотного
Все рабочие механизмы крана питаются от электрического привода на постоянном токе. Это обеспечивает низкие посадочные скорости и помогает им регулировать скорость от нулевой до максимальной.
Транспортировка крана требует наличия специального транспортного оборудования. При перевозке по автомобильным дорогам с использованием седельного тягача, конструкцию крана разбирают на узлы, а при железнодорожных перевозках секции башенно-стрелового оборудования укладываются друг на друга без разборки крана по опорно-поворотному устройству. Оригинальный принцип агрегатного построения дает МКГС-100 преимущество перед другими кранами тяжелого класса, связанных с демонтажем и транспортировкой оборудования предельных размеров.
Цены на аренду гусеничных кранов
Преимущества гусеничного крана МКГС-100
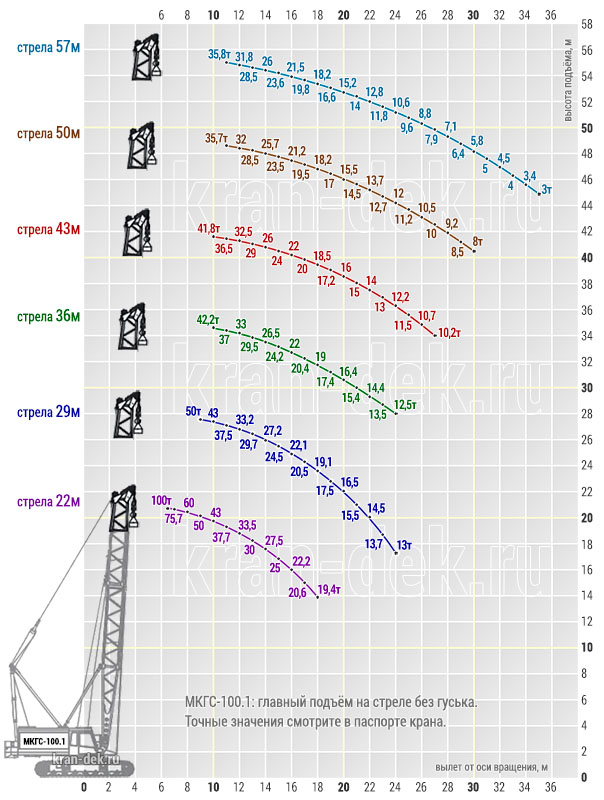
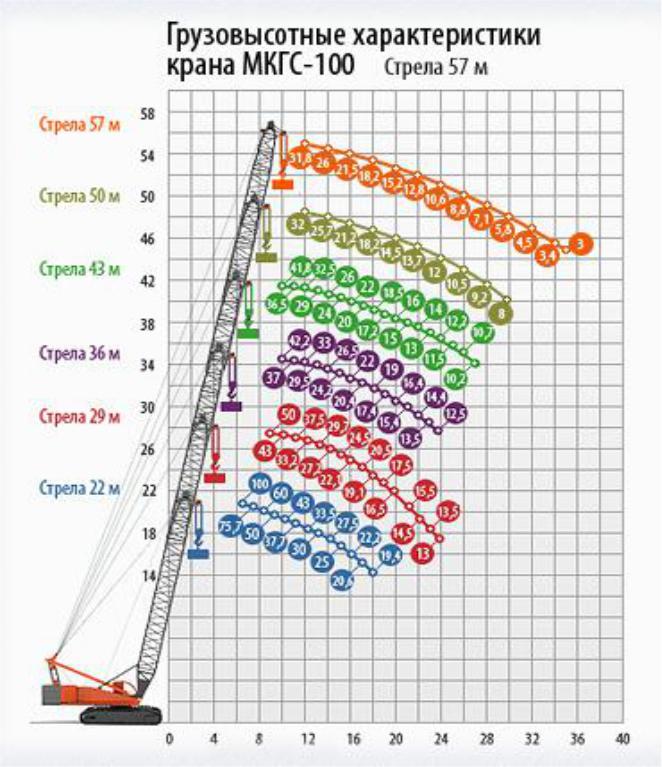
- Грузоподъемность. Максимальная грузоподъемность крана МКГС-100 при стреле длиной 22 м равна 100 т, при этом вылет стрелы равен 6 м, а высота подъема 20,7 м.

- Габариты. Ширина ходового устройства крана составляет 7000 мм с учетом ширины гусеничной ленты 1250 мм, длина — 9100 мм, а высота — 4250. Дорожный просвет составляет 550 мм. Рабочая масса крана равна 131,5 тоннам, включая массу контргруза, весом 21,8т и массу ходового и опорно-поворотного устройств массой 57,9 т. Скорость передвижения крана при средней нагрузке на грунт, равной 0,95 кгс/см¬2 составляет 0,5 км/час. Кран МКГС-100 способен преодолевать уклон пути не превышающий 15 град.
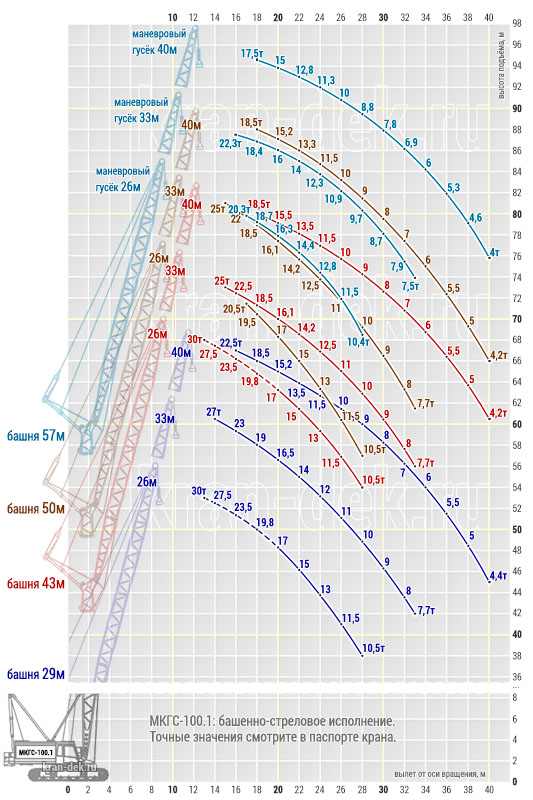
- Стреловое оборудование. Длина основной стрелы равна 22 м, однако за счет вставок 7 и 14 метров, она может увеличиться до 57 м. При этом наибольшая высота груза составит 96 м. Стрелы длиной 22, 29, 36, 43 и 50 метров, оснащенные 12-метровым жестким гуськом могут поднимать груз как на основном, так и на вспомогательном крюке. Подъем на максимальную высоту 96 м осуществляется с помощью башенно-стрелового оборудования длиной 57 м и гуськом 40 м при вылете стрелы 17 м. Вес груза в таком случае составляет 18,5 т.
- Электрооборудование. На кран МКГС-100 установлен двигатель марки ЯМЗ-236
мощностью 180 л.с. и числом оборотов, равным 1450 об/мин и 100-киловаттный генератор. Кран также приспособлен к получению энергии от внешней сети. Для этого необходим трехфазный ток напряжением 380В с частотой 50 Гц либо напряжением 440В частотой 60 Гц.
Грузовые характеристики МКГС-100.1
Заинтересовались? — Отправляйте заявку!
Наш электронный адрес [email protected] — обсчитаем оперативно!
Гусеничные краны МКГС-32, МКГС-100, МКГС-125.01, МКГС-300 в наличии и под заказ от Компании СПЕЦНЕФТЕМАШ
Гусеничный кран МКГС-125.01Технические характеристики МКГС-125.01 Грузоподъемность, т 125 Вылет при наибольшей грузоподъемности, м 6,5 Наибольшая высота башни, м 64 Габариты стрелы основной стрелы, м 22 наибольшей стрелы, м 57 Вставка для удлинения стрелы (башни), м 7; 14 Длина гуська для стрелы, м 12 Длина гуська для башни, м 26; 33; 40 Наибольшая высота подъема груза, м 95 Масса груза, поднимаемого на наибольшую высоту, т 19,5 Частота вращения поворотной платформы, мин 0,04 — 0,4 Скорость наматывания основного подъема на барабан, м/мин 36 Скорость передвижения, км/ч 0,6 Масса в полностью снаряженном состоянии (с основной стрелой), т 142 Давление на грунт, кПа/кгс/см2 102 / 1,02 Кратность запасовки каната при наибольшей грузоподъемности 11 при наибольшей высоте подъема груза 2 Кран гусеничный монтажный МКГС-125.
 Максимальный вылет стрелы от оси крана составляет 44 метра. Гусек крана МКГС-125.01 секционный различной длины (12, 26, 33, 40м) увеличивает подстреловое пространство, а, следовательно, и высоту подъёма груза. Кран МКГС-125.01 с электрическим приводом может работать от электрической сети или от собственной дизель-электростанции мощностью 100 кВт.
Безопасное проведение работ во всем диапазоне грузовых характеристик МКГС-125.01 обеспечивает микропроцессорный ограничитель грузоподъемности. Кроме того, микропроцессорный ограничитель грузоподъемности защищает кран МКГС-125.01 от перегрузки и опрокидывания по умолчанию. При выполнении подъемных операций в вблизи линии электропередач безопасность обеспечивает встроенный модуль защиты от опасного напряжения. Микропроцессорный ограничитель грузоподъемности оборудуется встроенным прибором фиксации характеристик, так называемым «черным ящиком», автоматически регистрирующим наиболее важные параметры.
Дизель-электростанция ДЭС-100, поставляемая с краном МКГС-125.
Максимальный вылет стрелы от оси крана составляет 44 метра. Гусек крана МКГС-125.01 секционный различной длины (12, 26, 33, 40м) увеличивает подстреловое пространство, а, следовательно, и высоту подъёма груза. Кран МКГС-125.01 с электрическим приводом может работать от электрической сети или от собственной дизель-электростанции мощностью 100 кВт.
Безопасное проведение работ во всем диапазоне грузовых характеристик МКГС-125.01 обеспечивает микропроцессорный ограничитель грузоподъемности. Кроме того, микропроцессорный ограничитель грузоподъемности защищает кран МКГС-125.01 от перегрузки и опрокидывания по умолчанию. При выполнении подъемных операций в вблизи линии электропередач безопасность обеспечивает встроенный модуль защиты от опасного напряжения. Микропроцессорный ограничитель грузоподъемности оборудуется встроенным прибором фиксации характеристик, так называемым «черным ящиком», автоматически регистрирующим наиболее важные параметры.
Дизель-электростанция ДЭС-100, поставляемая с краном МКГС-125.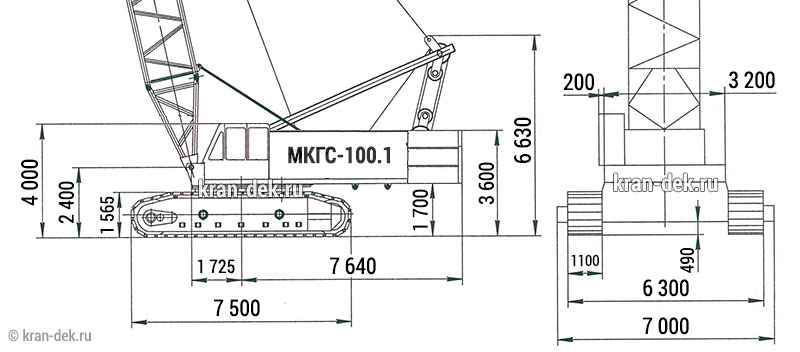
 01.
Продажа МКГС-125.01 осуществляется в нашей компании. Менеджеры компании ознакомят вас с техническими характеристиками выбранной модели гусеничных кранов Ульяновского механического завода №2, а также представят всю необходимую информацию. Если вы решили купить монтажный кран МКГС-125.01, свяжитесь с нами по контактным телефонам, указанным на нашем сайте в разделе «Контак.. далее
01.
Продажа МКГС-125.01 осуществляется в нашей компании. Менеджеры компании ознакомят вас с техническими характеристиками выбранной модели гусеничных кранов Ульяновского механического завода №2, а также представят всю необходимую информацию. Если вы решили купить монтажный кран МКГС-125.01, свяжитесь с нами по контактным телефонам, указанным на нашем сайте в разделе «Контак.. далееАренда гусеничного крана МКГС 100 тонн
Характеристики при башенно-стреловом исполнении
Высота башни,H
| Длина стрелы,l | Вылет, R | Высота подъема,h | грузоподъемность, тн |
|---|---|---|---|---|
| 29 | 26 | 13÷28 | 53÷38 | 31÷10,5 |
| 43 | 33 | 15÷33 | 73÷56 | 25÷7,7 |
| 50 | 40 | 17÷40 | 89÷66 | 19,5÷4,2 |
Характеристики при стреловом исполнении
| Длина стрелы, L | Вылет, R | Высота подъема, H | Грузоподъемность, тн |
|---|---|---|---|
| 22 | 6÷18 | 23÷16 | 100÷13,5 |
| 36 | 9÷24 | 37÷29 | 47,2÷5,5 |
| 50 | 9÷28 | 51÷44 | 46,6÷2,5 |
Технические характеристики
| Характеристики | Значение |
|---|---|
| Грузоподъемность, т | 100 |
| Вылет при наибольшей грузоподъемности, м | 6,5 |
| Длина основной стрелы, м | 22 |
| Наибольшая длина стрелы, м | 57 |
| Наибольшая высота башни, м | 57 |
| Длина вставок для удлинения, м | 7;14 |
| Длина гуська для стрелы, м | 12 |
| Длина гуська для башни, м | 26; 33; 40 |
| Наибольшая высота подъема груза, м | 95 |
| Груз поднимаемый на наибольшую высоту, м | 19,5 |
| Частота вращения поворотной платформы, об/мин | 0,04-0,4 |
| Скорость наматывания каната основного подъема на барабан, м/мин | 36 |
| Скорость передвижения, км/ч | 0,5 |
| Масса в полностью снаряженном состоянии, с основной стрелой, т | 130 |
| Среднее давление на грунт без груза, кг/см2 | 0,953 |
| Кратность запасовки каната при наибольшей грузоподъемности | 9 |
| Кратность запасовки каната при наибольшей высоте подъема груза | 2 |
| Статический коэффициент устойчивости относительно ребра опрокидывания | 71,40% |
Гусеничный кран МКГС-100 — это монтажный кран на гусеничном ходу.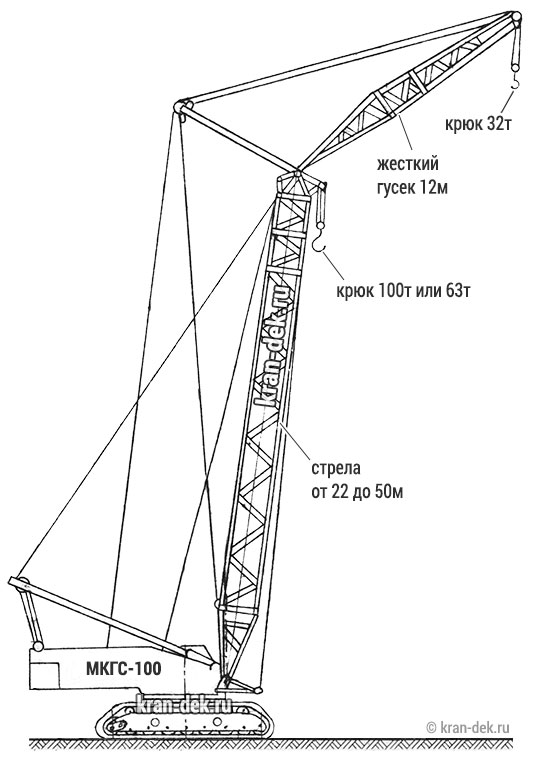
Кран используется в монтаже крупноблочного и тяжеловесного оборудования, металлических и других конструкций при строительстве заводов и электростанций.
МКГС-100 имеет электрический привод всех рабочих механизмов и это обеспечивает гибкое регулирование скоростей. Кран может работать от собственной дизель-станции, а также от внешней сети напряжением 380 В. Дизель-электрический агрегат МКГС-100 выполнен в отдельном модуле и не монтируется на кран.
Аренда гусеничного крана МКГС-100 — технические характеристики, заказать кран в Москве по низкой цене от компании «Монтаж-строй»
Технические характеристики
| Тип: | Автокраны |
| Грузоподъемность максимальная, т | 100 |
| Вылет при максимальной грузоподъемности, м | 6.5 |
| Длина основной стрелы, м | 22 |
| Наибольшая длина стрелы, м | 57 |
| Наибольшая высота башни, м | 57 |
| Длина вставок для удлинения стрелы, м | 7; 14 |
| Длина гуська для стрелы, м | 12 |
| Длина гуська для башни, м | 26; 33; 40 |
| Наибольшая высота подъема груза, м | 95 |
| Груз, поднимаемый на наибольшую высоту, т | 19. 5 5 |
| Частота вращения поворотной части, об/мин | 0.04 — 0.4 |
| Скорость наматывания каната основного подъема на барабан, м/мин | 36 |
| Скорость передвижения, км/ч | 0.5 |
| Масса в полностью снаряженном состоянии (с основной стрелой), т | 130 |
| Среднее давление на грунт без груза, кг/см² | 0.953 |
| Кратность запасовки каната при наибольшей грузоподъемности | 9 |
| Кратность запасовки каната при наибольшей высоте подъема груза | 2 |
| Статический коэффициент устойчивости относительно ребра опрокидования | 71.4% |
Цена аренды:
| от 8150 руб/час |
| от 65000 руб/смену |
Гусеничный кран МКГС-100. 1 предназначен для проведения крупномасштабных погрузочно-разгрузочных, ремонтных и монтажных работ в промышленном, дорожном и гражданском строительстве. Спецтехника позволяет перемещать до 100 тонн груза. Модель МКГС-100.1 отличается повышенной морозостойкостью, что обеспечивает возможность эксплуатации при температуре до -40 °С. Компания «Монтаж-строй» предлагает услуги аренды автокрана с возможностью доставки оборудования на строительный объект. При необходимости наши специалисты могут выполнять техобслуживание крана в течение всего периода аренды.
1 предназначен для проведения крупномасштабных погрузочно-разгрузочных, ремонтных и монтажных работ в промышленном, дорожном и гражданском строительстве. Спецтехника позволяет перемещать до 100 тонн груза. Модель МКГС-100.1 отличается повышенной морозостойкостью, что обеспечивает возможность эксплуатации при температуре до -40 °С. Компания «Монтаж-строй» предлагает услуги аренды автокрана с возможностью доставки оборудования на строительный объект. При необходимости наши специалисты могут выполнять техобслуживание крана в течение всего периода аренды.
Другая спецтехника из раздела: Гусеничные автокраны
Ульяновец МКГС-100
Гусеничный кран МКГС-100 — мощный, неприхотливый, работающий в самых сложных условиях. Грузовысотные характеристики значительно превышают зарубежные аналоги. Максимальная длина стрелы 57 м, длина маневрового гуська 40 м. Кран МКГС-100 прост в монтаже и перевозится по автодорогам при помощи специального транспортного устройства, поставляемого в комплекте.
| Двигатель | |
| Тип двигателя | дизельный |
| Основные характеристики | |
| Грузоподъемность крана, кг | 100000 |
| Масса крана, кг | 130000 |
| Полное название | Гусеничный кран Ульяновец МКГС-100 |
| Размеры | |
| Вылет стрелы при макс. грузоподъемности, м | 6.5 |
| Топливная система | |
| Максимальная скорость, км/ч | 0. 5 5 |
| Характеристика автокрана | |
| Длина гуська, м | стрелы-12. башни-26; 33; 40 |
| Длина стрелы, м | 22-57 |
| Максимальная высота подъема груза, м | 95 |
| Частота вращения, об/мин. | 0.04-0.4 |
| № п/п | Наименование машин и механизмов | Тип, модель | Кол-во | Основные технические характеристики |
| 1 | Кран гусеничный | МКГС-100 | 2 | грузоподъемность 100 тонн |
| 2 | Кран гусеничный | СКГ-63/100 | 2 | грузоподъемность 63 тонн |
| 3 | Кран гусеничный | МКГ-40 | 2 | грузоподъемность 40 тонн |
| 4 | Кран гусеничный | МКГ-25БР | 3 | грузоподъемность 25 тонн |
| 5 | Кран гусеничный | РДК-25 | 2 | грузоподъемность 25 тонн |
| 6 | Кран гусеничный | МКГ-16 | 3 | грузоподъемность 16 тонн |
| 7 | Кран автомобильный | LTM-1100/2 | 1 | грузоподъемность 100 тонн |
| 8 | Кран автомобильный | КС-6476 | 1 | грузоподъемность 50 тонн |
| 9 | Кран автомобильный | КС-55713-1 | 2 | грузоподъемность 25 тонн |
| 10 | Кран автомобильный | КС-45717 | 1 | грузоподъемность 25 тонн |
| 11 | Кран автомобильный | КС-4572 | 2 | грузоподъемность 16 тонн |
| 12 | Кран автомобильный | КС-35715 | 1 | грузоподъемность 16 тонн |
| 13 | Кран автомобильный | КС-35715-1 | 2 | грузоподъемность 15 тонн |
| 14 | Кран автомобильный | КС-3574 | 1 | грузоподъемность 14 тонн |
| 15 | Кран козловой | КК-12,5 | 1 | грузоподъемность 12,5 тонн |
| 16 | Кран погрузчик | МКРС-300П | 1 | грузоподъемность 12,5 тонн |
| 17 | Кран погрузчик | КП-300 | 2 | грузоподъемность 10 тонн |
| 18 | Кран башенный | КБ-403 | 1 | грузоподъемность 8 тонн |
| 19 | Кран башенный | КБ-100 | 1 | грузоподъемность 5 тонн |
| 20 | Автовышка | АГП-18 | 1 | высота 18 м |
| 21 | Автомобиль тягач | МАН-26480 | 1 | грузоподъемность 19 тонн |
| 22 | Автомобиль тягач | МАН-33440 | 1 | грузоподъемность 26 тонн |
| 23 | Автомобиль тягач | МАН-33480 | 1 | грузоподъемность 26 тонн |
| 24 | Автомобиль тягач | МАЗ-642208 | 2 | грузоподъемность 17 тонн |
| 25 | Автомобиль тягач | КАМАЗ-54112 | 2 | |
| 26 | Автомобиль тягач | КРАЗ-258 | 2 | |
| 27 | Автомобиль бортовой | КАМАЗ-53200 | 3 | |
| 28 | Автомобиль бортовой | КРАЗ-257 | 1 | грузоподъемность 12 тонн |
| 29 | Автомобиль самосвал | КРАЗ-6510 | 3 | |
| 30 | Автомобиль самосвал | КАМАЗ-5511 | 1 | |
| 31 | Полуприцеп самосвал | LANGENDORF 9539 | 1 | объем 34 куб. м. м. |
| 32 | Полуприцеп тентованный | ШМИДТ | 1 | длина 13,6 м |
| 33 | Полуприцеп самосвал | MELLER | 1 | грузоподъемность 35 тонн |
| 34 | Полуприцеп цементовоз | Сеспель | 1 | грузоподъемность 35 тонн |
| 35 | Полуприцеп бортовой | 1 | длина 12 м | |
| 36 | Полуприцеп бортовой | АБ-8М | 1 | длина 13,5 м |
| 37 | Полуприцеп панелевоз | 1 | грузоподъемность 12 тонн | |
| 38 | Спец.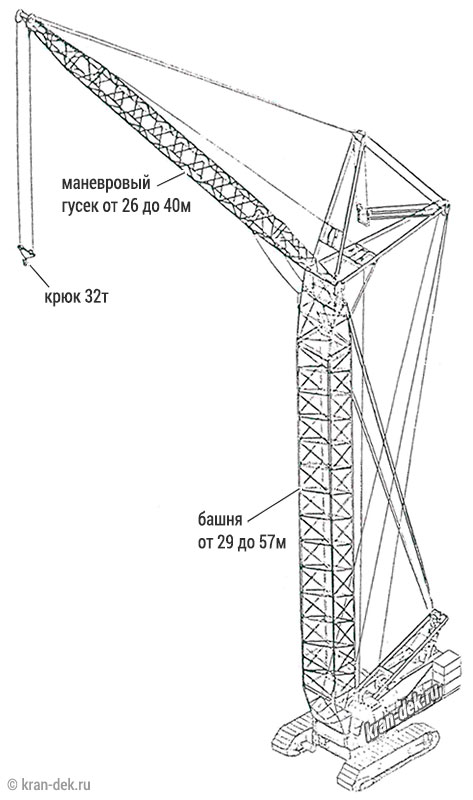 трейлер трейлер |
1 | грузоподъемность 50 тонн | |
| 39 | Трейлер | 1 | грузоподъемность 50 тонн | |
| 40 | Трейлер | 1 | грузоподъемность 25 тонн | |
| 41 | Автомобиль с крановоманипуляторной установкой | MAHLE | 2 | грузоподъемность 5 тонн |
| 42 | Экскаватор | Hundai | 1 | объем 1 куб. м. |
| 43 | Экскаватор | EK-18 | 1 | объем 1 куб. м. м. |
| 44 | Гидромолот | Дельта | 1 | |
| 45 | Трубоукладчик | ТГ-123 | 2 |
Аренда гусеничного крана МКГС 100 1 − Парк техники SWTrans
Аренда гусеничного крана МКГС 100.1
грузоподъемность 100 тонн
Гусеничный монтажный кран МКГС-100.1 грузоподъемностью 100 тонн состоит из ходового устройства, поворотной части, лебедки, механизма вращения платформы, генераторной группы, дизель-электрического агрегата, монтажной стойки, кабины оператора, опорно-поворотного устройства, стрелового оборудования, башенно-стрелового оборудования, крюковых обойм.
Гусеничный кран МКГС 100.1 мощный и неприхотливый, готов к работе в самых сложных условиях строительных площадок. Взяв в аренду МКГС 100. 1 вы получите в работу простой в монтаже и эксплуатации кран. Доставку арендованной техники компания SWTRANS выполняет собственными автотранспортными средствами.
1 вы получите в работу простой в монтаже и эксплуатации кран. Доставку арендованной техники компания SWTRANS выполняет собственными автотранспортными средствами.
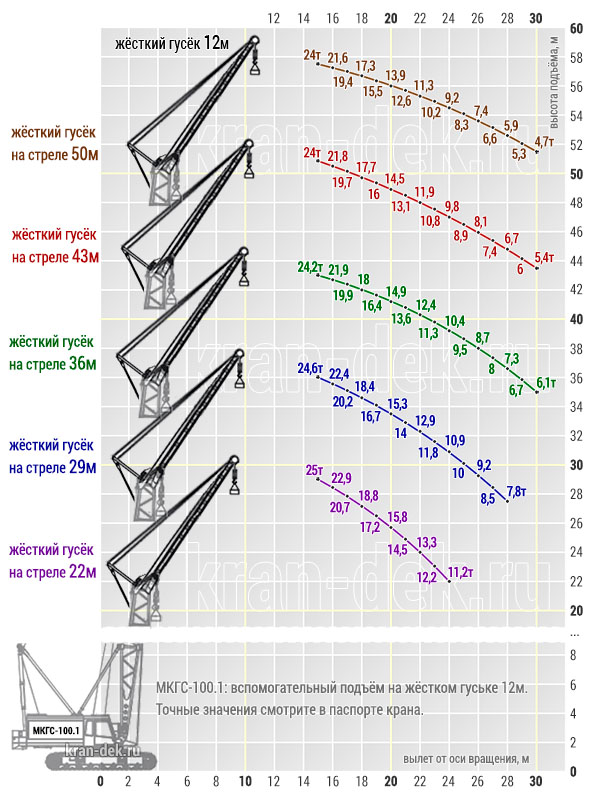
Стрела длиной 22, 29, 36,43, 50 м с жестким гуськом длиной 12 м имеет преимущество в том, что груз поднимается или на основном крюке, или на вспомогательном.
Максимальный груз Q = 100 т поднимается на высоту Н = 20,7 м при вылете l = 6,5 м на основном крюке, груз Q = 25 т поднимается на высоту Н = 29 м при вылете l= 15 м на вспомогательном крюке на стреле длиной L = 22 м.
На максимальную высоту Н= 48,7 м при вылете l = 9 м поднимается груз Q = 47 т на основном крюке и на высоту Н = 57,5 м при вылете l = 15 м поднимается груз Q = 24 т на вспомогательном крюке на стреле длиной L = 50 м.
Башня (башенно-стреловое исполнение) длиной 29, 43, 50, 57м с гуськом длиной 26, 33, 40м работает с грузом на большой высоте: поднимает груз Q =19,5 т на высоту Н = 95 м при вылете l = 17 м на башне 57 м с гуськом 40 м.
Технические характеристики крана МКГС-100.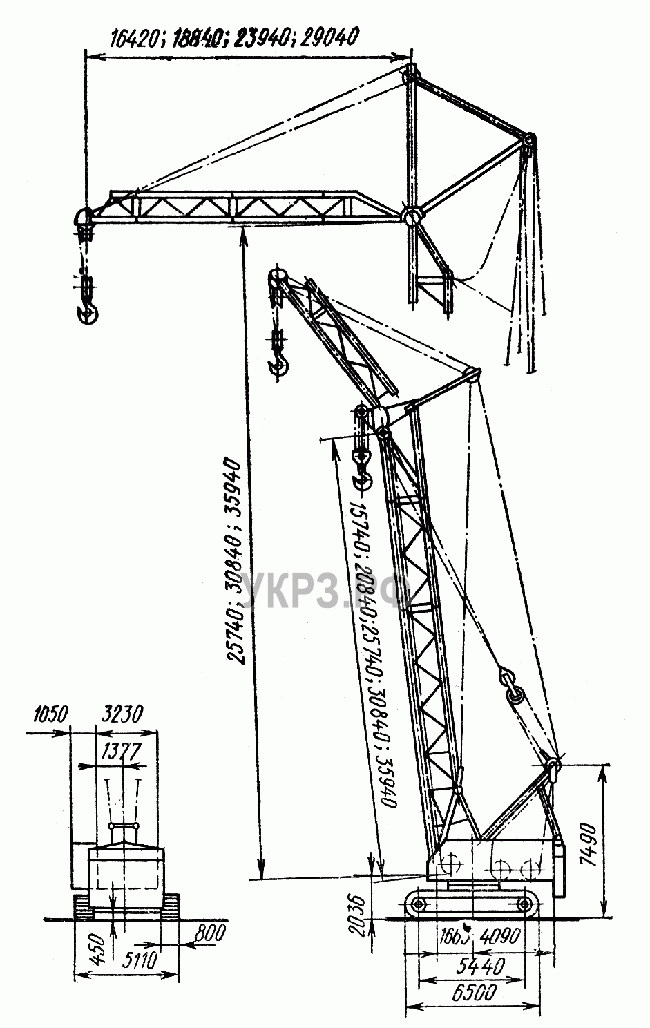 1
1
Грузоподъемность — 100 т
Вылет при наибольшей грузоподъемности — 6,5 м
Длина основной стрелы — 22 м
Наибольшая длина стрелы — 57 м
Наибольшая высота башни — 57 м
Длина вставок для удлинения стрелы — 7 м
Длина гуська для стрелы — 12м
Длина гуська для башни — 26, 30 м
Наибольшая высота подъема груза — 95 м
Груз поднимаемый на наибольшую высоту — 17,5, м
Скорость передвижения — 0,8-8,33 км/ч
Масса в полностью снаряженном состоянии — 130 т
Среднее давление на грунт без груза — 0,953 кг/см
Длительная аренда гусеничного крана МКГС 100.1 — выгодное предложение для строительных компаний!
Вы можете оставить заявку на аренду гусеничного крана МКГС 100.1 непосредственно на странице или обратившись в нашу компанию по телефонам, указанным на сайте. Если у Вас остались вопросы по комплектации, наши менеджеры предоставят Вам полную информацию об условиях аренды крана и другой спецтехники.
обеспечивает независимую температурную чувствительность термочувствительных микрогелей и позволяет переключать плазмонное связывание, а также контролировать поглощение и высвобождение
Введена новая гибридная система ДНК-микрогель с двойной термочувствительностью, обеспечивающая ковалентное связывание одноцепочечной ДНК (оцДНК) с термореактивными микрогелями (мкГс). Пространственное распределение сайтов связывания оцДНК было охарактеризовано с помощью трехмерной флуоресцентной микроскопии сверхвысокого разрешения.Функционализированные ДНК мкГ остаются термореактивными и могут поглощать меченную красителем комплементарную оцДНК, которая может снова высвобождаться путем преодоления температуры дегибридизации ДНК независимо от объемного фазового перехода (VPT) мкГ. То же самое верно и для нанообъектов, представленных плазмонными наночастицами золота (AuNP), глубина проникновения которых была визуализирована с помощью ПЭМ-томографии и 3D-реконструкции и которые демонстрируют усиленную плазмонную связь в свернутом состоянии мкГс и, таким образом, становятся переключаемыми. Напротив, если оцДНК была поглощена только неспецифическими взаимодействиями, , то есть , в нефункционализированные мкГ, ее высвобождение не зависит от температуры и может быть вызвано только увеличением концентрации соли. Таким образом, встроенная оцДНК представляет сайты связывания с высокой степенью отбора, определяемые их числом оснований и последовательностью, что делает VPT, определяемую составом мкГ, и обратимое поглощение и высвобождение, обеспечиваемое посредством программируемой гибридизации ДНК, являются независимыми характеристиками.Сочетание типично высокой биосовместимости и сохраняющейся набухаемости и проницаемости открывает перспективы для новых фундаментальных исследований, а также для потенциальных применений в биологических средах.
Напротив, если оцДНК была поглощена только неспецифическими взаимодействиями, , то есть , в нефункционализированные мкГ, ее высвобождение не зависит от температуры и может быть вызвано только увеличением концентрации соли. Таким образом, встроенная оцДНК представляет сайты связывания с высокой степенью отбора, определяемые их числом оснований и последовательностью, что делает VPT, определяемую составом мкГ, и обратимое поглощение и высвобождение, обеспечиваемое посредством программируемой гибридизации ДНК, являются независимыми характеристиками.Сочетание типично высокой биосовместимости и сохраняющейся набухаемости и проницаемости открывает перспективы для новых фундаментальных исследований, а также для потенциальных применений в биологических средах.
У вас есть доступ к этой статье
Подождите, пока мы загрузим ваш контент. ..
Что-то пошло не так. Попробуй еще раз?
..
Что-то пошло не так. Попробуй еще раз?(PDF) Методология двухуровневой балансировки нагрузки с критерием выбора потребителей на основе кластеризации для размещения коммутационных устройств в микросетях
Ссылки
1.Халид, М .; Умер Акрам; Шафик, С., Оптимальное планирование нескольких распределенных генерирующих устройств и хранилища
в активных распределительных сетях, IEEE Access 2018, 6, 55234 — 55244.
2. Григорас Г.; Иванов, О .; Neagu, B.C .; Прагма К. Стратегии, основанные на интеллектуальном измерении, для повышения энергоэффективности
Эффективность микросетей. В архитектуре микросетей, методы управления и защиты; Mahdavi Tabatabaei N. ,
,
Kabalci E., Bizon N., Eds .; Springer-Verlag, Берлин, Германия, 2020 г., стр.463-490.
3. Liu, B .; Meng, K .; Dong, Z .; Вонг, П. К. С .; Ли, X. Балансировка нагрузки в низковольтной распределительной сети с помощью
Phase Reconfiguration: эффективный подход, основанный на чувствительности, в IEEE Trans. на Pow. Deliv. 2020, Early
Доступ doi: 10.1109 / TPWRD.2020.3022061.
4. Haq, S.U .; Ариф, Б .; Хан, А .; Ахмед, Дж. Автоматическая трехфазная система балансировки нагрузки с использованием быстрого переключающего реле
в трехфазной системе распределения.Труды Международной конференции по энергетике,
Energy and Smart Grid (ICPESG), Мирпур, Пакистан, 2018, стр. 1-6, DOI: 10.1109 / ICPESG.2018.8384514.
5. Narayanan, K. N .; Умананд, Л. Новый маршрутизатор с активной фазой для динамической балансировки нагрузки в трехфазной микросети
. Труды Азиатско-Тихоокеанской конференции по энергетике и энергетике IEEE PES 2017
(APPEEC), Бангалор, Индия, 2017 г. , стр. 1-5, DOI: 10.1109 / APPEEC.2017.8308922.
, стр. 1-5, DOI: 10.1109 / APPEEC.2017.8308922.
6.Homaee, O .; Najafi, A .; Дехганян, М .; Attar, M .; Фалаги, Х. Практический подход к распределению
балансировки сетевой нагрузки путем оптимального переключения фаз однофазных клиентов с использованием дискретного генетического алгоритма.
Внутр. Пер. по системе электроэнергетики. 2019, 29 (5), e2834.
7. Liu, Y.-W .; Rau, S.-H .; Wu, C.-J .; Ли, В.-Дж. Повышение качества электроэнергии за счет использования усовершенствованной компенсации реактивной мощности
. IEEE Trans. Ind. Appl. 2018, 54, 18–24.
8. Калесар, Б.М. Заказчики переключаются между фазами для снижения потерь с учетом дневного профиля нагрузки
в интеллектуальной сети. В материалах семинара CIRED 2016, Хельсинки, Финляндия, 14–15 июня 2016 г .; С.
1–4.
9. Jianguo, Z .; Qiuye, S .; Huaguang Z .; Янь, З. Балансировка нагрузки и компенсация реактивной мощности на основе шунтовой компенсации конденсаторных батарей
в распределительных сетях низкого напряжения, Труды 31-й китайской конференции по управлению
, Хэфэй, Китай, 2012 г. , стр.6681-6686.
, стр.6681-6686.
10. Arias, J .; Calle, M .; Turizo, D .; Guerrero, J .; Кандело-Бесерра, Дж. Э. Историческая балансировка нагрузки в распределительных системах
, использующих алгоритм перехода и границы. Энергия 2019, 12, 1219.
11. Granada Echeverri, M .; Gallego Rendón, R.A .; Лопес Лезама, Дж. М. Планирование оптимального баланса фаз для снижения потерь
в системах распределения с использованием специализированного генетического алгоритма. Ingeniería y Ciencia 2012, 8 (15),
121–140.
12.Аль-Харсан, И. Х. Новая стратегия балансировки нагрузки с переключением фаз, основанная на метаэвристическом алгоритме
MOGWO, J. Mech. Продолжить. Математика. Sci. 2020, 15 (2), 84–102.
13. Mahendran, G .; Говиндараджу, К. Алгоритм опыления цветов для балансировки фаз распределительной системы
с учетом переменного спроса. Microprocessors and Microsystems 2020, 74, 103008.
14. Saffar, A .; Hooshmand, R .; Ходабахшян, А. Новая нечеткая оптимальная реконфигурация распределительных систем
Новая нечеткая оптимальная реконфигурация распределительных систем
для снижения потерь и балансировки нагрузки с использованием алгоритма поиска муравьиной колонии.Приложение. Soft Comp.
2011, 11 (5), 4021-4028.
15. Hooshmand, R .; Солтани, С. Х. Одновременная оптимизация фазового баланса и реконфигурации в распределительных сетях
с использованием алгоритма BF – NM. Int. J. of Elec. Pow. & En. Syst. 2012, 41 (1), 76-86.
16. Олек, Б .; Вежбовски, М. Балансировка местной энергии и вспомогательные услуги в низковольтных сетях с распределенной генерацией
, накоплением энергии и активными нагрузками. IEEE Trans.на Инд. Электр. 2014, 62 (4), 2499-2508.
17. Кихавани, М. Р .; Гаджизаде, А .; Шахириния, А. Координация зарядки и балансировка нагрузки подключаемых электрических транспортных средств
в несбалансированных низковольтных распределительных системах, IET Gen., Transm. & Дистриб. 2020, 14 (3), 389-399,
DOI: 10. 1049 / iet-gtd.2019.0397.
1049 / iet-gtd.2019.0397.
18. Weckx, S .; Дризен Дж. Балансировка нагрузки с помощью зарядных устройств для электромобилей и фотоэлектрических инверторов в несбалансированных распределительных сетях.
IEEE Trans. на Sust. En.2015, 6 (2), 635-643.
19. Евзельман, М .; Ур Рехман, М. М .; Hathaway, K .; Zane, R .; Costinett D .; Максимович Д. Система активной балансировки
для электромобилей со встроенной низковольтной шиной. IEEE Trans. на Pow. Электр. 2015, 31 (11),
7887-7895, DOI: 10.1109 / TPEL.2015.2513432.
20. Faessler, B .; Schuler, M .; Preißinger, M .; Кепплингер, П. Системы хранения аккумуляторов как мера балансировки энергосистемы
в низковольтных распределительных сетях с распределенной генерацией.Энергия 2017, 10, 2161.
Препринты (www.preprints.org) | НЕ РЕЗУЛЬТАТЫ | Опубликовано: 9 декабря 2020 г. doi: 10.20944 / preprints202012.0226.v1
Антибактериальные применения нанокомпозитов α-Fe 2 O 3 / Co 3 O 4 и изучение их структурных, оптических, магнитных и цитотоксических характеристик
Abu-Zied BM , Soliman SA (2009) Разложение закиси азота на катализаторах MCO 3 –Co 3 O 4 (M = Ca, Sr, Ba). Catal Lett 132: 299–310
Catal Lett 132: 299–310
Статья Google Scholar
Араха М., Пал С., Самантаррай Д., Паниграхи Т.К., Маллик Б.К., Праманик К., Маллик Б., Джа С. (2015) Антимикробная активность наночастиц оксида железа при модуляции интерфейса наночастицы-бактерии.Sci Rep 5: 14813. https://doi.org/10.1038/srep14813
Артикул Google Scholar
Aruguete DM, Bojeong K, Michael FH, Yanjun M, Yingwen C, Andy H, Jie L, Amy P (2013) Антимикробная нанотехнология: ее потенциал для эффективного управления микробной лекарственной устойчивостью и значение для исследовательских потребностей в микробиологических исследованиях. нанотоксикология. Воздействие экологических процессов 15: 93–102
Статья Google Scholar
Basnet P, Larsen GK, Jadeja RP, Hung YC, Zhao Y (2013) α-Fe 2 O 3 наноколонки и наностержни, изготовленные методом электронно-лучевого испарения для фотокаталитических и антимикробных применений в видимом свете. Интерфейсы приложения ACS Mater 5 (6): 2085–2095. https://doi.org/10.1021/am303017c
Интерфейсы приложения ACS Mater 5 (6): 2085–2095. https://doi.org/10.1021/am303017c
Артикул Google Scholar
Bauer AW, Kirby WM, Sherris JC, Turck M (1966) Тестирование чувствительности к антибиотикам стандартным методом с одним диском. Am J Clin Pathol 45 (4): 493–496
Статья Google Scholar
Блечер К., Насир А., Фридман А. (2011) Растущая роль нанотехнологий в борьбе с инфекционными заболеваниями.Вирулентность 2: 395–401
Статья Google Scholar
Brayner R, Dahoumane SA, Yepremian C, Djediat C, Meyer M, Coute A, Fiévet F (2010) Наночастицы ZnO: синтез, характеристика и экотоксикологические исследования. Langmuir 26: 6522–6528
Статья Google Scholar
Cesar I, Kay A, Gonzalez Martinez JA, Grätzel M (2006) Прозрачные тонкопленочные фотоаноды Fe 2 O 3 для эффективного расщепления воды солнечным светом: направляемый наноструктурами эффект легирования кремнием. J Am Chem Soc 128: 4582–4583
J Am Chem Soc 128: 4582–4583
Статья Google Scholar
Chambers HF, DeLeo FR (2009) Волны устойчивости: золотистый стафилококк в эпоху антибиотиков. Nat Rev Microbiol 7: 629–641
Статья Google Scholar
Chen L, Yang X, Chen J, Liu J, Wu H, Zhan H, Liang C, Wu M (2010) Непрерывная настройка формы и спектроскопия нанокристаллов гематита. Inorg Chem 49: 8411–8420
Статья Google Scholar
Chen T, Wang R, Xu LQ, Neoh KG, Kang ET (2012) Магнитные наночастицы, функционализированные карбоксиметил хитозаном, для разрушения биопленок Staphylococcus aureus и Escherichia coli .Ind Eng Chem Res 51: 13164–13172
Статья Google Scholar
Chipara M, Ibrahim E, Yust B, Padilla D, Chipara DM (2015) Наночастицы и бактерии. Журнал Nanomed Res 2 (3): 00033. https://doi.org/10.15406/jnmr.2015.02.00033
https://doi.org/10.15406/jnmr.2015.02.00033
Артикул Google Scholar
Dasari TP, Pathakoti K, Hwang HM (2013) Определение механизма фотоиндуцированной токсичности выбранных наночастиц оксидов металлов (ZnO, CuO, Co 3 O 4 и TiO 2 ) на E.coli бактерий. J Environ Sci 25: 882–888
Статья Google Scholar
Дэвис Дж., Дэвис Д. (2010) Истоки и эволюция устойчивости к антибиотикам. Microbiol Mol Biol Rev 74: 417–433
Статья Google Scholar
Денизот Ф., Ланг Р. (1986) Быстрый колориметрический анализ роста и выживаемости клеток: модификации процедуры окрашивания тетразолием, обеспечивающие повышенную чувствительность и надежность.J Immunol Methods 89 (2): 271–277
Статья Google Scholar
Foster HA, Ditta IB, Varghese S, Steele A (2011) Фотокаталитическая дезинфекция с использованием диоксида титана: спектр и механизм антимикробной активности. Appl Microbiol Biotechnol 90: 1847–1868
Appl Microbiol Biotechnol 90: 1847–1868
Статья Google Scholar
Fu G, Vary PS, Lin CT (2005) Anatase TiO 2 Нанокомпозиты для антимикробных покрытий.J Phys Chem B 109: 8889–8898
Статья Google Scholar
Ghosh T, Dash SK, Chakraborty P, Guha A, Kawaguchi K, Roy S, Chattopadhyay T, Das D (2014) Получение антиферромагнитных наночастиц Co 3 O 4 наночастиц из двух различных прекурсоров пиролитическим методом: антимикробная активность in vitro. RSC Adv 4: 15022–15029
Статья Google Scholar
Хаджипур М.Дж., Фромм К.М., Ашкарран А.А., Хименес де Аберастури Д., Ларраменди IRD, Рохо Т., Серпушан В., Парак В.Дж., Махмуди М. (2012) Антибактериальные свойства наночастиц.Trends Biotechnol 30: 499–511
Статья Google Scholar
Худ М. И., Скаар Е.П. (2012) Пищевой иммунитет: переходные металлы на границе патоген-хозяин. Nat Rev Microbiol 10: 525–537
И., Скаар Е.П. (2012) Пищевой иммунитет: переходные металлы на границе патоген-хозяин. Nat Rev Microbiol 10: 525–537
Статья Google Scholar
Huh AJ, Kwon YJ (2011) «Наноантибиотики»: новая парадигма лечения инфекционных заболеваний с использованием наноматериалов в эпоху устойчивости к антибиотикам.J Control Release 156: 128–145
Артикул Google Scholar
Jayaraman R (2009) Устойчивость к антибиотикам: обзор механизмов и смена парадигмы. Curr Sci 96: 1475–1484
Google Scholar
Хан С., Ансари А.А., Хан А.А., Ахмад Р., Аль-Обайд О., Аль-Каттан В. (2015) Оценка in vitro противоопухолевой и антибактериальной активности наночастиц оксида кобальта.Журнал J Biol Inorg Chem 20 (8): 1319–1326. https://doi.org/10.1007/s00775-015-1310-2
Артикул Google Scholar
Кохански М. А., Дуайер Д.Д., Хайете Б., Лоуренс Калифорния, Коллинз Дж. Дж. (2007) Общий механизм гибели клеток, вызванный бактерицидными антибиотиками. Cell 130: 797–810
А., Дуайер Д.Д., Хайете Б., Лоуренс Калифорния, Коллинз Дж. Дж. (2007) Общий механизм гибели клеток, вызванный бактерицидными антибиотиками. Cell 130: 797–810
Статья Google Scholar
Kong H, Song J, Jang J (2010) Одностадийное производство магнитных наночастиц γ-Fe 2 O 3 / полиданина с использованием химической окислительной полимеризации in situ и их антибактериальных свойств.Chem Commun 46: 6735–6737
Статья Google Scholar
Леви С.Б., Маршалл Б. (2004) Устойчивость к антибактериальным препаратам во всем мире: причины, проблемы и ответы. Nat Med 10: S122 – S129. https://doi.org/10.1038/nm1145
Артикул Google Scholar
Лю Дж., Аругете Д.М., Мураяма М., Хочелла М.Ф. (2009) Влияние размера и агрегации на реакционную способность экологически и промышленно значимого наноматериала (PbS). Environ Sci Technol 43: 8178–8183
Environ Sci Technol 43: 8178–8183
Статья Google Scholar
Махлуф С., Дрор Р., Ницан Ю., Абрамович Ю., Елинек Р., Геданкен А. (2005) Синтез нанокристаллического MgO с помощью микроволнового излучения и его использование в качестве бактериоцида. Adv Funct Mater 15: 1708–1715
Статья Google Scholar
Маринкович-Станоевич З.В., Ромчевич Н., Стоянович Б.Д. (2007) Спектроскопическое исследование шпинели ZnCr 2 O 4 4 , полученной из механически активированных смесей ZnO – Cr 2 O 3 .J Eur Ceram Soc 27: 903–907
Статья Google Scholar
Moritz M, Geszke-Moritz M (2013) Новейшие достижения в синтезе, иммобилизации и практическом применении антибактериальных наночастиц. Chem Eng J 228: 596–613
Статья Google Scholar
Мосманн Т. (1983) Быстрый колориметрический анализ клеточного роста и выживаемости: применение для анализов пролиферации и цитотоксичности.J Immunol Methods 65 (1-2): 55–63
(1983) Быстрый колориметрический анализ клеточного роста и выживаемости: применение для анализов пролиферации и цитотоксичности.J Immunol Methods 65 (1-2): 55–63
Статья Google Scholar
Nor YA, Zhou L, Meka AK, Xu C, Niu Y, Zhang H, Mitter N, Mahony D, Yu C (2016) Разработка полых наносфер из оксида железа для улучшения антимикробных свойств: понимание цитотоксического происхождения в богатых органическими веществами среда. Adv Funct Mater 26: 5408–5418
Статья Google Scholar
Nussbaum FV, Brands M, Hinzen B, Weigand S, Häbich D (2006) Антибактериальные натуральные продукты в медицинской химии — исход или возрождение? Angew Chem Int Ed 45: 5072–5129
Статья Google Scholar
Pareta RA, Taylor E, Webster TJ (2008) Повышенная плотность остеобластов в присутствии новых магнитных наночастиц, покрытых фосфатом кальция. Нанотехнологии 19: 265101. https://doi.org/10.1088/0957-4484/19/26/265101
Нанотехнологии 19: 265101. https://doi.org/10.1088/0957-4484/19/26/265101
Артикул Google Scholar
Parham S, Wicaksono DHB, Bagherbaigi S, Lee SL, Nur H (2016) Антимикробная обработка наночастиц различных оксидов металлов: критический обзор. J Chin Chem Soc 63: 385–393
Статья Google Scholar
Pelgrift RY, Friedman AJ (2013) Нанотехнологии как терапевтический инструмент для борьбы с микробной резистентностью.Adv Drug Deliv Rev 65: 1803–1815
Артикул Google Scholar
Rafi MM, Ahmed KSZ, Nazeer KP, Kumar DS, Thamilselvan M (2015) Синтез, характеристика и магнитные свойства наночастиц гематита (α-Fe 2 O 3 ) на полисахаридных матрицах и их антибактериальная активность. Appl Nanosci 5: 515–520. https://doi.org/10.1007/s13204-014-0344-z
Артикул Google Scholar
Ramana CV, Massot M, Julien CM (2005) XPS и рамановское спектроскопическое исследование LiMn 2 O 4 шпинелей. Surf Interface Anal 37: 412–416
Surf Interface Anal 37: 412–416
Статья Google Scholar
Rudramurthy GR, Swamy MK, Sinniah UR, Ghasemzadeh A (2016) Наночастицы: альтернативы лекарственно-устойчивым патогенным микробам. Молекулы 21: 836. https://doi.org/10.3390/molecules21070836
Артикул Google Scholar
Руфус А., Сриджу Н., Филип Д. (2016) Синтез наночастиц биогенного гематита (α-Fe 2 O 3 ) для применения в антибактериальных и наножидкостных целях.RSC Adv 6: 94206–94217. https://doi.org/10.1039/C6RA20240C
Артикул Google Scholar
Шарма П., Рана Д.С., Ахмад У., Кумар Р., Неги К., Чаухан М.С., Чаухан С. (2016) Нанокубцы из оксида железа для фотокаталитического разложения и антимикробных применений. Nanosci Nanotechnol Lett 8: 1014–1019
Статья Google Scholar
Stankic S, Suman S, Haque F, Vidic J (2016) Чистые и многометаллические наночастицы оксидов металлов: синтез, антибактериальные и цитотоксические свойства. J Nanobiotechnol 14:73. https://doi.org/10.1186/s12951-016-0225-6
J Nanobiotechnol 14:73. https://doi.org/10.1186/s12951-016-0225-6
Артикул Google Scholar
Touati D (2000) Железо и окислительный стресс у бактерий. Arch Biochem Biophys 373: 1–6
Статья Google Scholar
Тран Н., Мир А., Маллик Д., Синха А., Наяр С., Вебстер Т.Дж. (2010) Бактерицидное действие наночастиц оксида железа на Staphylococcus aureus .Int J Nanomed 5: 277–283
Google Scholar
Ван Х., Уильямс Р.Л., Доэрти П.Дж., Уильямс Д.Ф. (1994) Оценка цитотоксичности кевлара и карбида кремния с помощью анализа МТТ. J Mater Sci 5 (6–7): 441–445
Google Scholar
Wang Z, Lee YH, Wu B, Horst A, Kang Y, Tang YJ, Chen DR (2010) Антимикробная активность аэрозольных наночастиц оксида переходных металлов.Chemosphere 80: 525–529
Статья Google Scholar
Вайнберг ED (1999) Нагрузка железом и наблюдение за болезнями. Emerg Infect Dis 5: 346–352
Emerg Infect Dis 5: 346–352
Статья Google Scholar
Xia Y (2008) Наноматериалы в работе в биомедицинских исследованиях. Nat Mater 7: 758–760
Статья Google Scholar
Zborowski M, Chalmers JJ (2007) Магнитное разделение ячеек, 1-е изд.Эльзевир, Амстердам
Google Scholar
Zhang H, Chen B, Banfield JF (2010) Влияние размера частиц и pH на растворение наночастиц. J Phys Chem C 114: 14876–14884
Статья Google Scholar
Zhang Q, Lambert G, Liao D, Kim H, Robin K, Tung CK, Pourmand N, Austin RH (2011) Ускорение появления бактериальной устойчивости к антибиотикам в связанных микросредах.Science 333 (6050): 1764–1767. https://doi.org/10.1126/science.1208747
Артикул Google Scholar
Каррагинан — обзор | Темы ScienceDirect
23.
 3.3 Системы на основе каррагинана
3.3 Системы на основе каррагинана Каррагинан — линейный сульфатированный полисахарид, экстрагированный из внеклеточного матрикса (ВКМ) красных морских водорослей семейства Rhodophyceae. Химическая структура каррагинана состоит из звеньев β-d-галактозы и 3,6-ангидро-α-d-галактозы, объединенных гликозидными связями и содержащих сложноэфирные сульфатные группы, частично сульфатированные [7,9,39].С коммерческой точки зрения каррагинаны делятся на три семейства в зависимости от количества и положения сульфатных групп: ι (йота), κ (каппа) и λ (лямбда), содержащих одну, две и три сульфатные группы на дисахарид. Первые два являются желирующими агентами, а третий — загустителем, который чаще всего используется в биомедицине [7,9,39]. Недавно была продемонстрирована полезность каррагинанов в системах доставки лекарств для лечения рака. Чен и др. синтезировал наночастицу каррагенанового олигосахарида-золота (Car-AuNPs), которая использовалась в качестве системы доставки для доставки эпирубицина, лекарственного средства, используемого для химиотерапии, с триггером pH [104].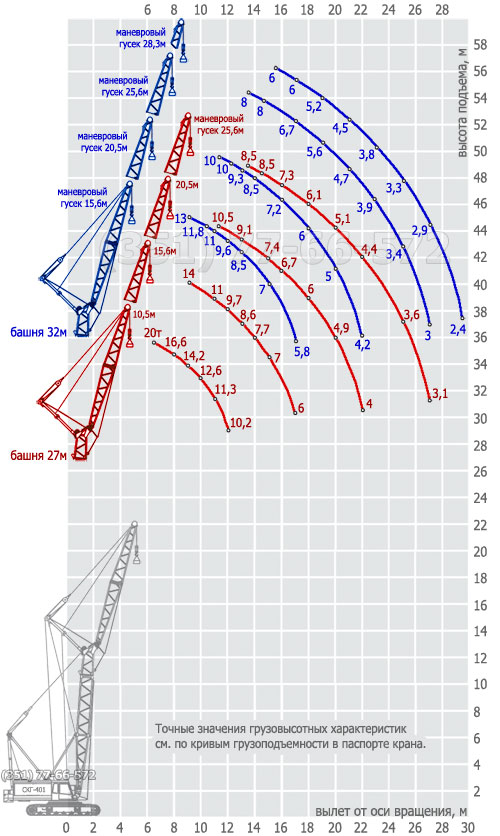 Цитотоксичность Car-AuNP, эпирубицина (EPI) и нагруженных эпирубицином Car-AuNP (EPI-Car-AuNP) оценивали с использованием нормальных (HUVEC) и клеток рака печени (HepG2). Car-AuNP не влияет на жизнеспособность нормальных клеток, подтверждая, что эти NP обладают хорошей биосовместимостью. EPI-Car-AuNP проявляли более низкую цитотоксичность по сравнению с нормальными клетками, чем свободные EPI. И EPI, и EPI-Car-AuNP демонстрируют дозозависимое снижение жизнеспособности клеток против клеток HepG2, и ингибирование EPI-Car-AuNPs было более значительным, чем свободный EPI при той же дозе.Кроме того, EPI-Car-AuNPs индуцировали апоптоз клеток посредством остановки клеточного цикла в фазе G2 / M в клетках HepG2.
Цитотоксичность Car-AuNP, эпирубицина (EPI) и нагруженных эпирубицином Car-AuNP (EPI-Car-AuNP) оценивали с использованием нормальных (HUVEC) и клеток рака печени (HepG2). Car-AuNP не влияет на жизнеспособность нормальных клеток, подтверждая, что эти NP обладают хорошей биосовместимостью. EPI-Car-AuNP проявляли более низкую цитотоксичность по сравнению с нормальными клетками, чем свободные EPI. И EPI, и EPI-Car-AuNP демонстрируют дозозависимое снижение жизнеспособности клеток против клеток HepG2, и ингибирование EPI-Car-AuNPs было более значительным, чем свободный EPI при той же дозе.Кроме того, EPI-Car-AuNPs индуцировали апоптоз клеток посредством остановки клеточного цикла в фазе G2 / M в клетках HepG2.
В другом исследовании гибридные наноструктурированные микрочастицы (hNPs) на основе карбоната кальция и λ-каррагинана (λ-Car), дериватизированные фолиевой кислотой (FA), были разработаны для доставки DOX [105]. Цитотоксический эффект чНЧ FA-λ-Car с или без DOX оценивали на клеточной линии, полученной из остеобластной остеосаркомы человека MG-63. HNP FA-λ-Car-DOX значительно снижали жизнеспособность клеток MG-63, в то время как FA-λ-Car — нет.Кроме того, цитотоксичность FA-λ-Car-DOX была аналогична цитотоксичности, индуцированной супернатантами hNP FA-λ-Car-DOX. Совсем недавно для улучшения доставки DOX был синтезирован GO-привитой наноноситель κ-каррагинана, конъюгированный с биотином (GO-κ-Car-biotin) [106]. Нагруженный DOX наноноситель GO-κ-кар-биотин значительно снижал жизнеспособность клеток линии клеток карциномы шейки матки (HeLa) в зависимости от концентрации, но наблюдалось только небольшое снижение жизнеспособности линии клеток дермальных фибробластов человека.Клетки, обработанные наноносителем GO-κ-кар-биотина, нагруженным DOX, показали неправильную форму ядра, сжатие хроматина, разбросанные ядерные гранулы, что предполагает ядерную фрагментацию. Клеточная пролиферация значительно снизилась, а утечка ЛДГ значительно увеличилась в зависимости от концентрации за счет нагруженного DOX GO-κ-кар-биотина.
HNP FA-λ-Car-DOX значительно снижали жизнеспособность клеток MG-63, в то время как FA-λ-Car — нет.Кроме того, цитотоксичность FA-λ-Car-DOX была аналогична цитотоксичности, индуцированной супернатантами hNP FA-λ-Car-DOX. Совсем недавно для улучшения доставки DOX был синтезирован GO-привитой наноноситель κ-каррагинана, конъюгированный с биотином (GO-κ-Car-biotin) [106]. Нагруженный DOX наноноситель GO-κ-кар-биотин значительно снижал жизнеспособность клеток линии клеток карциномы шейки матки (HeLa) в зависимости от концентрации, но наблюдалось только небольшое снижение жизнеспособности линии клеток дермальных фибробластов человека.Клетки, обработанные наноносителем GO-κ-кар-биотина, нагруженным DOX, показали неправильную форму ядра, сжатие хроматина, разбросанные ядерные гранулы, что предполагает ядерную фрагментацию. Клеточная пролиферация значительно снизилась, а утечка ЛДГ значительно увеличилась в зависимости от концентрации за счет нагруженного DOX GO-κ-кар-биотина.
Наноструктурированные гибридные носители липид-каррагинан (NLCC) были использованы для контролируемой доставки химиотерапевтического агента митоксантрон гидрохлорида (MTO) [107]. Относительное клеточное поглощение MTO-NLCC было значительно выше, чем для раствора MTO (MTO-Sol) в клетках MCF-7 / MX, сверхэкспрессирующих белок устойчивости к раку молочной железы (BCRP). Соответственно, цитотоксичность MTO была значительно увеличена в MTO-NLCC по сравнению с MTO-Sol. Напротив, после инкубации с пустыми NLCC не было обнаружено значительного ингибирования роста, что указывает на то, что NLCC были нетоксичными в качестве носителя. In vivo Фармакокинетика MTO-Sol и MTO-NLCC у крыс продемонстрировала, что очевидная биодоступность группы MTO-NLCC значительно увеличилась по сравнению с группой MTO-Sol.
Относительное клеточное поглощение MTO-NLCC было значительно выше, чем для раствора MTO (MTO-Sol) в клетках MCF-7 / MX, сверхэкспрессирующих белок устойчивости к раку молочной железы (BCRP). Соответственно, цитотоксичность MTO была значительно увеличена в MTO-NLCC по сравнению с MTO-Sol. Напротив, после инкубации с пустыми NLCC не было обнаружено значительного ингибирования роста, что указывает на то, что NLCC были нетоксичными в качестве носителя. In vivo Фармакокинетика MTO-Sol и MTO-NLCC у крыс продемонстрировала, что очевидная биодоступность группы MTO-NLCC значительно увеличилась по сравнению с группой MTO-Sol.
Куркумин продемонстрировал антипролиферативный эффект при множественных формах рака и преодолел его плохую растворимость, Sathuvan et al. приготовили нагруженные куркумином гранулы κ-каррагинана (κ-Car-Cur) для доставки лекарств против клеток рака легких (A549) [108]. Тесты на клетках in vitro показали, что композиты κ-Car-Cur вызывают более высокую цитотоксичность в отношении раковых клеток, чем свободный куркумин. Кроме того, производство ROS увеличилось, а потенциал митохондриальной мембраны снизился во время лечения κ-Car-Cur, что предполагает, что эта система запускает клеточный апоптоз в раковых клетках A549.
Кроме того, производство ROS увеличилось, а потенциал митохондриальной мембраны снизился во время лечения κ-Car-Cur, что предполагает, что эта система запускает клеточный апоптоз в раковых клетках A549.
Использование имеющегося в продаже набора для экстракции ДНК для получения высококачественной геномной ДНК человека, подходящей для ПЦР и генотипирования, из 11-летних ватных тампонов, насыщенных слюной | Примечания к исследованию BMC
участников
Семейное исследование штата Вермонт представляет собой коллекцию образцов из 207 семей, состоящих из 783 человек. 167 семей были частью выборки с СДВГ, причем по одному члену каждой семьи признали пробандом для этого расстройства. 40 семей были включены в контрольные семьи без диагнозов DSM-IV.
Сбор образцов
Участников попросили воздержаться от еды и питья за 1 час до сбора слюны. Каждому пациенту было предложено поместить стандартный кусок хлопковой марли размером 2 × 2 дюйма в щечную область щеки на 3 минуты. Насыщенный слюной ватный тампон удаляли, скатывали, чтобы он соответствовал пробирке для сбора, и хранили при -20 ° C до тех пор, пока не была извлечена геномная ДНК. Все данные собраны и проанализированы с одобрения этического комитета UVM COM IRB.
Насыщенный слюной ватный тампон удаляли, скатывали, чтобы он соответствовал пробирке для сбора, и хранили при -20 ° C до тех пор, пока не была извлечена геномная ДНК. Все данные собраны и проанализированы с одобрения этического комитета UVM COM IRB.
Экстракция и количественное определение ДНК
Все экстракции ДНК были выполнены в Институте генетики поведения человека Avera.ДНК экстрагировали из пропитанных слюной ватных тампонов с использованием метода очистки на колонке. Свернутые тампоны для слюны имели размеры приблизительно 5 см × 1,5 см, и ДНК экстрагировали из буккальных клеток с использованием протокола большого объема QIAamp DNA Blood Midi Kit (Qiagen) в соответствии с инструкциями производителя с небольшими изменениями. Пыж инкубировали при 70 ° C в смеси буфера протеаза / лизис (200 мкл протеазы Qiagen / 2,4 мл буфера AL) в течение 30 минут в конической пробирке на 15 мл (Fisher Scientific).Лизат отделяли от слюны с использованием центробежной силы путем помещения слюны в цилиндр шприца на 5 мл (Becton Dickson), помещенного в коническую пробирку на 15 мл, и центрифугировали в течение 10 минут при 12000 об / мин. Пыж отбрасывали, к лизату добавляли 2 мл абсолютного этанола и перемешивали пробирку путем энергичного встряхивания (встряхивания). Примерно половину смеси лизат / этанол переносили в колонку Qiagen Midi, помещенную в чистую коническую пробирку на 15 мл. Колонку центрифугировали при 1850 × g в течение 3 минут.Фильтрат отбрасывали, а оставшуюся смесь лизат / этанол наносили на ту же колонку и центрифугировали при 1850 × g в течение 3 минут. Колонку промывали 2 мл буфера AW1 и центрифугировали при 3220 × g в течение 2 минут. Колонку промывали второй раз 2 мл буфера AW2 и центрифугировали при 3220 × g в течение 30 минут, чтобы гарантировать полное высыхание. ДНК элюировали из колонки в чистую коническую пробирку на 15 мл, добавляя в колонку 200 мкл буфера AE, инкубируя при комнатной температуре в течение 5 минут и центрифугируя при 3220 × g в течение 4 минут.Для максимального выхода ДНК проводили вторую элюцию, как описано выше, с получением общего объема приблизительно 400 мкл. Концентрацию и чистоту ДНК определяли с помощью УФ-спектрофотометрии (Nanodrop).
Пыж отбрасывали, к лизату добавляли 2 мл абсолютного этанола и перемешивали пробирку путем энергичного встряхивания (встряхивания). Примерно половину смеси лизат / этанол переносили в колонку Qiagen Midi, помещенную в чистую коническую пробирку на 15 мл. Колонку центрифугировали при 1850 × g в течение 3 минут.Фильтрат отбрасывали, а оставшуюся смесь лизат / этанол наносили на ту же колонку и центрифугировали при 1850 × g в течение 3 минут. Колонку промывали 2 мл буфера AW1 и центрифугировали при 3220 × g в течение 2 минут. Колонку промывали второй раз 2 мл буфера AW2 и центрифугировали при 3220 × g в течение 30 минут, чтобы гарантировать полное высыхание. ДНК элюировали из колонки в чистую коническую пробирку на 15 мл, добавляя в колонку 200 мкл буфера AE, инкубируя при комнатной температуре в течение 5 минут и центрифугируя при 3220 × g в течение 4 минут.Для максимального выхода ДНК проводили вторую элюцию, как описано выше, с получением общего объема приблизительно 400 мкл. Концентрацию и чистоту ДНК определяли с помощью УФ-спектрофотометрии (Nanodrop). Всю геномную ДНК либо разбавляли, либо концентрировали до конечной концентрации 50 нг / мкл. ДНК разводили в буфере с восстановленным EDTA (10 мМ Tris-HCL, 0,1 мМ EDTA, pH 8,0) и концентрировали с использованием устройств с центрифужными фильтрами Microcon YM-100 (Millipore) в соответствии с инструкциями производителя.
Всю геномную ДНК либо разбавляли, либо концентрировали до конечной концентрации 50 нг / мкл. ДНК разводили в буфере с восстановленным EDTA (10 мМ Tris-HCL, 0,1 мМ EDTA, pH 8,0) и концентрировали с использованием устройств с центрифужными фильтрами Microcon YM-100 (Millipore) в соответствии с инструкциями производителя.
ПЦР-амплификация
Качество ДНК, выделенной из слюны, оценивали с помощью ПЦР-амплификации полиморфной области, связанной с переносчиком серотонина (5HTTLPR).Продукты ПЦР визуализировали с помощью электрофореза в 2% агарозном геле или анализа фрагментов на генетическом анализаторе ABI 3130 (Applied Biosystems, Inc.). Последовательности праймеров для 5-HTTLPR были описаны ранее; прямой праймер (5′-ATGCCAGCACCTAACCCCTAATGT-3 ‘) и обратный праймер (5′-GGACCGCAAGGTGGGCGGGA-3’) [7]. При прогоне образцов на генетическом анализаторе 3130 для пометки продукта ПЦР для анализа фрагментов использовали флуоресцентно помеченный прямой праймер (6FAM, Applied Biosystems). Эта пара праймеров амплифицирует продукт из 419 пар оснований для аллеля с 16 повторами длинной (L) и продукт из 375 пар оснований для аллеля с 14 повторами короткого (S).Реакции ПЦР проводили с использованием основной смеси для ПЦР (Promega), содержащей конечную концентрацию 1,5 мМ MgCl 2 , 1 × реакционный буфер, 200 мкМ каждого dNTP, 40 нг очищенной геномной ДНК, 1,25 единиц ДНК-полимеразы Taq и 5 пмоль каждого праймера на 25 мкл реакции. Условия цикла ПЦР состояли из начальной денатурации при 95 ° C в течение 15 минут, 35 циклов, каждый из которых состоял из 30 секунд при 94 ° C, 30 секунд при 66 ° C и 40 секунд при 72 ° C. После последнего цикла удлинение продолжали в течение 15 мин при 72 ° C.Фрагменты S по сравнению с L были определены с использованием программного обеспечения GeneMapper версии 4.0 (ABI), или фрагменты были разделены на 2% агарозном геле с добавлением бромистого этидия (0,02%, Fisher).
Эта пара праймеров амплифицирует продукт из 419 пар оснований для аллеля с 16 повторами длинной (L) и продукт из 375 пар оснований для аллеля с 14 повторами короткого (S).Реакции ПЦР проводили с использованием основной смеси для ПЦР (Promega), содержащей конечную концентрацию 1,5 мМ MgCl 2 , 1 × реакционный буфер, 200 мкМ каждого dNTP, 40 нг очищенной геномной ДНК, 1,25 единиц ДНК-полимеразы Taq и 5 пмоль каждого праймера на 25 мкл реакции. Условия цикла ПЦР состояли из начальной денатурации при 95 ° C в течение 15 минут, 35 циклов, каждый из которых состоял из 30 секунд при 94 ° C, 30 секунд при 66 ° C и 40 секунд при 72 ° C. После последнего цикла удлинение продолжали в течение 15 мин при 72 ° C.Фрагменты S по сравнению с L были определены с использованием программного обеспечения GeneMapper версии 4.0 (ABI), или фрагменты были разделены на 2% агарозном геле с добавлением бромистого этидия (0,02%, Fisher).
Европа PMC
Cell Rep.2018 23 октября; 25 (4): 921–933. e5.
e5.
Катерина Гкирциманаки
1 Институт молекулярной биологии и биотехнологий Ираклион, Греция
2 Лаборатория аутоиммунитета и воспаления медицинского факультета Критского университета, Ираклион, Греция
Элени Кабрани
2 Лаборатория аутоиммунитета и воспаления медицинского факультета Критского университета, Гераклион Греция
Димитра Николери
2 Лаборатория аутоиммунитета и воспаления, Медицинский факультет, Университет Крита, Ираклион, Греция
Александр Полизос
905 30 4 Фонд биомедицинских исследований Афинской академии, Афины, Греция
Афанасиос Бланас
2 Лаборатория аутоиммунитета и воспаления, медицинский факультет Критского университета, Ираклион, Греция
Prodromos Sidiropoulos
2 Лаборатория аутоиммунитета и воспаления, Медицинский факультет Критского университета, Ираклион, Греция3 Кафедра ревматологии, Университетская больница Ираклиона, Медицинский факультет, Критский университет, Ираклион, Греция
Антонис Макригианнакис
905 Кафедра акушерства и гинекологии, Медицинский факультет, Университет Крита, Ираклион, Греция
Джордж Берциас
2 Лаборатория аутоиммунитета и воспаления, Медицинский факультет, Университет Крита, Ираклион, Греция
3 Ревматология, Университетская клиника Ираклиона, Медицинский факультет, U университет Крита, Ираклион, Греция
Димитриос Т.
 Бумпас
Бумпас5 4-й медицинский факультет, Университетская больница Аттикон, Национальный университет Каподистрии, Афины, Греция, и медицинская школа, Университет Кипра, Никосия, Кипр
6 Лаборатория иммунной регуляции и толерантности, аутоиммунитета и воспаления, Фонд биомедицинских исследований Афинской академии, Афины, Греция
Panayotis Verginis
6 Лаборатория иммунной регуляции и толерантности, аутоиммунитета и воспаления, Фонд биомедицинских исследований Афинской академии, Афины, Греция
1 Институт кафедры молекулярной биологии и биотехнологии, Фонд исследований и технологий, Ираклион, Греция
2 Лаборатория аутоиммунитета и воспаления, Медицинский факультет, Университет Крита, Ираклион, Греция
3 Отделение ревматологии, Университетская больница Ираклиона , Медицинский факультет, Университет Крита, Геракли on, Греция
4 Фонд биомедицинских исследований Афинской академии, Афины, Греция
5 4-й медицинский факультет, больница Университета Аттикон, Национальный университет Каподистрии, Афины, Греция, и медицинский факультет Кипрского университета, Никосия, Кипр
6 Лаборатория иммунной регуляции и толерантности, аутоиммунитета и воспаления, Фонд биомедицинских исследований Афинской академии, Афины, Греция
7 Кафедра акушерства и гинекологии, Медицинская школа, Университет Крита, Ираклион , Греция
8 Эти авторы внесли равный вклад
9 Текущий адрес: Иммунная регуляция и рак, Центр молекулярной медицины Макса Дельбрюка, Берлин 13125, Германия
10 Контактное лицо
Получено 1 сентября 2017 г . ; Пересмотрено 25 июня 2018 г .; Принята в печать 31 августа 2018 г.
; Пересмотрено 25 июня 2018 г .; Принята в печать 31 августа 2018 г.
Это статья в открытом доступе под лицензией CC BY-NC-ND (http://creativecommons.org/licenses/by-nc-nd/4.0/).
Эта статья цитируется в других статьях в PMC.Сопутствующие данные
- Дополнительные материалы
Документ S1. Рисунки S1 – S6 и таблицы S2 и S4
Таблица S1. Клинические записи пациентов с СКВ, относящиеся к рисунку 1
Таблица S3. RNA-Seq DEG, относящиеся к фиг. 6A. Таблица, содержащая 380 дифференциально экспрессируемых генов (DEG) между пациентом и группой здоровых людей.
Документ S2. Статья плюс дополнительная информация
Резюме
Интерферон α (IFNα) является быстрым и эффективным организатором защиты организма от нуклеиновых кислот, но при хроническом течении становится мощным медиатором аутоиммунитета. Устойчивая передача сигналов IFNα связана с патогенезом системной красной волчанки (СКВ), неизлечимого аутоиммунного заболевания, характеризующегося аберрантным восприятием собственной ДНК, которое достигает кульминации в патологии, опосредованной аутоантителами. IFNα инструктирует дифференцировку моноцитов в аутовоспалительные дендритные клетки (DC), чем потенцирует выживание и рост аутореактивных лимфоцитов, но молекулярный механизм, соединяющий тревогу стерильного IFNα-опасности с адаптивной реакцией против собственной ДНК, остается неуловимым.Здесь мы демонстрируем IFNα-опосредованную дерегуляцию митохондриального метаболизма и нарушение аутофагической деградации, что приводит к цитозольному накоплению мтДНК, которая воспринимается через стимулятор генов интерферона (STING), чтобы способствовать индукции аутовоспалительных DC. Идентификация мтДНК как клеточно-автономного усилителя передачи сигналов IFNα подчеркивает важность эффективного рециклинга митохондрий в поддержании периферической толерантности. Антиоксидантное лечение и метаболическое восстановление деградации аутолизосом становятся мишенями для лекарств при СКВ и других патологиях, связанных с IFNα.
IFNα инструктирует дифференцировку моноцитов в аутовоспалительные дендритные клетки (DC), чем потенцирует выживание и рост аутореактивных лимфоцитов, но молекулярный механизм, соединяющий тревогу стерильного IFNα-опасности с адаптивной реакцией против собственной ДНК, остается неуловимым.Здесь мы демонстрируем IFNα-опосредованную дерегуляцию митохондриального метаболизма и нарушение аутофагической деградации, что приводит к цитозольному накоплению мтДНК, которая воспринимается через стимулятор генов интерферона (STING), чтобы способствовать индукции аутовоспалительных DC. Идентификация мтДНК как клеточно-автономного усилителя передачи сигналов IFNα подчеркивает важность эффективного рециклинга митохондрий в поддержании периферической толерантности. Антиоксидантное лечение и метаболическое восстановление деградации аутолизосом становятся мишенями для лекарств при СКВ и других патологиях, связанных с IFNα.
Ключевые слова: аутоиммунитет, СКВ, лизосома, метаболизм
Графический реферат
Введение
Системная красная волчанка (СКВ) представляет собой сложное аутоиммунное воспалительное заболевание, характеризующееся множественными патофенотипами, в результате которых интерферон альдеринминология превращается в альфа-интерферон (IFNα) в форму. стойкие самонаправленные иммунные ответы против аутологичных нуклеиновых кислот. Чрезмерное и трудноизлечимое повреждение тканей, вызванное аутоантителами, особенно иммуноглобулином G (IgG) против двухцепочечной ДНК (дцДНК) или отложениями иммунных комплексов, влияет на различные органы, что приводит к увеличению заболеваемости и смертности (Weckerle et al., 2011). Полногеномные исследования ассоциации (GWAS) выявляют генетическую предрасположенность к активности IFN I типа при СКВ с семейной обмороженной волчанкой, рассматриваемой как интерферонопатия (Crow and Manel, 2015), а функциональные исследования подтверждают ее роль в нарушении периферической толерантности и развитии аутоиммунитета (Crow , 2014, Liao et al., 2016). В подтверждение этому у пациентов с гепатитом С и лейкемией, получавших IFNα, часто развивается волчаночный синдром с высокими титрами аутоантител против дцДНК.
стойкие самонаправленные иммунные ответы против аутологичных нуклеиновых кислот. Чрезмерное и трудноизлечимое повреждение тканей, вызванное аутоантителами, особенно иммуноглобулином G (IgG) против двухцепочечной ДНК (дцДНК) или отложениями иммунных комплексов, влияет на различные органы, что приводит к увеличению заболеваемости и смертности (Weckerle et al., 2011). Полногеномные исследования ассоциации (GWAS) выявляют генетическую предрасположенность к активности IFN I типа при СКВ с семейной обмороженной волчанкой, рассматриваемой как интерферонопатия (Crow and Manel, 2015), а функциональные исследования подтверждают ее роль в нарушении периферической толерантности и развитии аутоиммунитета (Crow , 2014, Liao et al., 2016). В подтверждение этому у пациентов с гепатитом С и лейкемией, получавших IFNα, часто развивается волчаночный синдром с высокими титрами аутоантител против дцДНК.
Передача сигналов IFNα и активация путей восприятия нуклеиновых кислот и клиренса взаимосвязаны (Shrivastav and Niewold, 2013), управляя противовирусным иммунитетом, таким как дифференцировка моноцитов в мощные антигенпредставляющие дендритные клетки (DC) с повышенной способностью управлять T и B клеточные ответы (Blanco et al. , 2001, Ding et al., 2006, Jegerlehner et al., 2007, Ma et al., 2013, Pascual et al., 2006, Rönnblom and Pascual, 2008). При SLE циркулирующие моноциты уже дифференцированы в DCs, их функция определяется IFNα (Blanco et al., 2001, Rönnblom and Pascual, 2008). Эти IFNα-DC демонстрируют повышенную экспрессию HLA-DR, CD86 и BAFF, активируют аутореактивные Т-клетки и способствуют размножению и дифференцировке аутореактивных В-клеток, таким образом объединяя врожденное восприятие опасности с адаптивным аутоиммунным ответом (Gujer et al., 2011). Однако молекулярный механизм (ы), управляющий IFNα-опосредованной аутореактивностью в антигенпрезентирующих клетках (APC) СКВ, остается неуловимым.
, 2001, Ding et al., 2006, Jegerlehner et al., 2007, Ma et al., 2013, Pascual et al., 2006, Rönnblom and Pascual, 2008). При SLE циркулирующие моноциты уже дифференцированы в DCs, их функция определяется IFNα (Blanco et al., 2001, Rönnblom and Pascual, 2008). Эти IFNα-DC демонстрируют повышенную экспрессию HLA-DR, CD86 и BAFF, активируют аутореактивные Т-клетки и способствуют размножению и дифференцировке аутореактивных В-клеток, таким образом объединяя врожденное восприятие опасности с адаптивным аутоиммунным ответом (Gujer et al., 2011). Однако молекулярный механизм (ы), управляющий IFNα-опосредованной аутореактивностью в антигенпрезентирующих клетках (APC) СКВ, остается неуловимым.
Аутофагия — это эволюционно законсервированный механизм переноса мембран, с помощью которого клетки доставляют цитоплазматические субстраты в лизосомы для их повторного использования для поддержания клеточного метаболического равновесия и гомеостаза (Klionsky et al., 2011). Появляющаяся литература предполагает участие аутофагии во внутриклеточном восприятии опасности, процессинге антигена и доставке для представления в APC (Alissafi et al. , 2017, Рённблом и Паскуаль, 2008). Более того, он регулирует активацию и дифференцировку APC посредством метаболической адаптации и поддержания митохондриального гомеостаза (Ravi et al., 2014, Zhang et al., 2012). Секвестрация и успешное очищение поврежденных митохондрий с помощью аутофагии (митофагии) подавляет накопление митохондриальных активных форм кислорода (mtROS) и предотвращает воспаление и образование аутоантигенов за счет внутриклеточного окисления (Minton, 2016). Многие антигены, участвующие в патологии СКВ, претерпели окислительную модификацию (дцДНК, ЛПВП, β2GPI, Ro), которая обнаруживает поразительную корреляцию с активностью заболевания и повреждением органов (Perl, 2013).Кроме того, нарушение аутофагического переваривания митохондриальных компонентов, таких как мтДНК, может привести к их цитозольной утечке и вызвать воспаление (Nakai et al., 2007). Высокоиммуногенные свойства мтДНК, неметилированной дцДНК бактериального происхождения (Pisetsky, 2016, Zhang et al., 2010), когда она не перерабатывается должным образом (Caielli et al.
, 2017, Рённблом и Паскуаль, 2008). Более того, он регулирует активацию и дифференцировку APC посредством метаболической адаптации и поддержания митохондриального гомеостаза (Ravi et al., 2014, Zhang et al., 2012). Секвестрация и успешное очищение поврежденных митохондрий с помощью аутофагии (митофагии) подавляет накопление митохондриальных активных форм кислорода (mtROS) и предотвращает воспаление и образование аутоантигенов за счет внутриклеточного окисления (Minton, 2016). Многие антигены, участвующие в патологии СКВ, претерпели окислительную модификацию (дцДНК, ЛПВП, β2GPI, Ro), которая обнаруживает поразительную корреляцию с активностью заболевания и повреждением органов (Perl, 2013).Кроме того, нарушение аутофагического переваривания митохондриальных компонентов, таких как мтДНК, может привести к их цитозольной утечке и вызвать воспаление (Nakai et al., 2007). Высокоиммуногенные свойства мтДНК, неметилированной дцДНК бактериального происхождения (Pisetsky, 2016, Zhang et al., 2010), когда она не перерабатывается должным образом (Caielli et al. , 2016, Oka et al., 2012), позволяют предположить что он может рассматриваться как сильный кандидат на управление аутореактивностью СКВ.
, 2016, Oka et al., 2012), позволяют предположить что он может рассматриваться как сильный кандидат на управление аутореактивностью СКВ.
В этом исследовании мы спросили, может ли нарушенная регуляция аутофагии быть вовлечена в IFNα-опосредованную иммуногенность моноцитов при СКВ и опосредовано ли это дефектами в их клеточной автономной рециркуляции, которая может привести к активации зондирования цитоплазматической ДНК.Наши результаты показывают, что характерная для волчанки подпись IFNα в моноцитах активных, нелеченных пациентов с СКВ связана с аберрантным митохондриальным метаболизмом, изменением лизосомного pH и дефектной аутофагической деградацией, которая приводит к накоплению мтДНК в цитоплазме. Стимулятор врожденных иммунных рецепторов генов интерферона (STING) воспринимает его и управляет дифференцировкой моноцитов в аутореактивные ДК, способствуя внутриклеточному ДНК-опосредованному, интерферон-зависимому иммунитету I типа. Расшифровка роли IFNα в автономной клеточной генерации и восприятии аутоантигенов в моноцитах волчанки предоставит нам важную информацию, которая может привести к лучшему нацеливанию лекарств на аутоиммунитет СКВ.
Результаты
Моноциты СКВ демонстрируют повышенное образование аутофагосом, но дефектную деградацию аутолизосом Чжоу и др., 2011). Чтобы понять регуляцию аутофагического потока в свежевыделенных моноцитах CD14
+ активных, нелеченных пациентов с СКВ (Таблица S1), мы отслеживали экспрессию и субклеточную локализацию необходимых компонентов аутофагии (Klionsky et al., 2016), а именно первую убиквитиноподобную систему конъюгации, которая необходима для удлинения фагофорной мембраны (Atg5-Atg12), вторая система конъюгации, необходимая для образования аутофагосом (PE-LC3B), секвестратор очищаемых субстратов ( SQSTM1 / P62), и кислотный отсек для переваривания субстратов аутофагии, лизосома (LAMP1). Как свидетельствует активация транскрипции ATG5, повышенное содержание липидированной формы LC3B и присутствие структур LC3 + P62 + LAMP1 + (A, 1B и 1D), задействуется аутофагия и усиливается образование аутофагосом. в моноцитах СКВ от активных пациентов. Примечательно, что неэффективная деградация, о чем свидетельствует накопление белка P62 в пузырьках LC3 + LAMP1 + в отсутствие его активации транскрипции (B-1D), указывает на то, что правильное разрешение автолизосом может быть затруднено. Нарушение функции аутолизосом может быть связано либо с нарушением слияния аутофагосом и лизосом, либо с неэффективным подкислением. Поскольку было обнаружено, что LAMP1 и LC3B puncta совместно локализуются в моноцитах СКВ (D), слияние аутофаго-лизосом, по-видимому, не затрагивается.Вместо этого было обнаружено, что кислотность лизосом снижена, поскольку Lysotracker Red-DND99 (LTR), флуорофор, связанный со слабым основанием, которое только частично протонируется при нейтральном pH, проявляет значительно более слабую интенсивность в моноцитах СКВ (E). Это изменение функционально влияет на лизосомальные ферменты, на что указывает снижение активности катепсина D (F). В совокупности эти результаты предполагают, что моноциты СКВ содержат пул аутофагосом, слитых с лизосомами, которые не подкислены должным образом для переваривания поврежденных внутриклеточных компонентов, что способствует накоплению неэффективно очищенных внутриклеточных отходов.
Примечательно, что неэффективная деградация, о чем свидетельствует накопление белка P62 в пузырьках LC3 + LAMP1 + в отсутствие его активации транскрипции (B-1D), указывает на то, что правильное разрешение автолизосом может быть затруднено. Нарушение функции аутолизосом может быть связано либо с нарушением слияния аутофагосом и лизосом, либо с неэффективным подкислением. Поскольку было обнаружено, что LAMP1 и LC3B puncta совместно локализуются в моноцитах СКВ (D), слияние аутофаго-лизосом, по-видимому, не затрагивается.Вместо этого было обнаружено, что кислотность лизосом снижена, поскольку Lysotracker Red-DND99 (LTR), флуорофор, связанный со слабым основанием, которое только частично протонируется при нейтральном pH, проявляет значительно более слабую интенсивность в моноцитах СКВ (E). Это изменение функционально влияет на лизосомальные ферменты, на что указывает снижение активности катепсина D (F). В совокупности эти результаты предполагают, что моноциты СКВ содержат пул аутофагосом, слитых с лизосомами, которые не подкислены должным образом для переваривания поврежденных внутриклеточных компонентов, что способствует накоплению неэффективно очищенных внутриклеточных отходов.
Передача сигналов IFNα нарушает завершение аутофагии и усиливает иммуногенный потенциал моноцитов
Чтобы выяснить, являются ли наблюдаемые дефекты завершения аутофагии присущими моноцитам СКВ или опосредованы растворимыми факторами воспалительной среды, мы культивировали моноциты CD14 + из здоровых доноры с сывороткой (10% об. / об.) от пациентов с активной СКВ (SLEDAI ≥8), активным ревматоидным артритом (РА) или здоровыми донорами и контролировали их аутофагический поток. Обработка здоровых моноцитов СКВ, но не сывороткой RA, воспроизводила фенотип моноцитов СКВ в отношении индукции аутофагии и дефектной деградации аутолизосом (рисунок S1; таблицы S1 и S2).Предыдущая работа показала, что моноциты, подвергшиеся воздействию сыворотки SLE, приобретают DC-подобный фенотип с повышенной способностью представлять антиген (Blanco et al., 2001). Чтобы выяснить, вовлечено ли аутолизосомное нарушение в этот процесс, мы метаболически модулировали аутофагический поток в здоровых моноцитах, культивируемых в присутствии сыворотки SLE, с использованием рапамицина, ингибитора механистической мишени передачи сигналов рапамицина (mTOR) и мощного усилителя образования аутофагосом. и деградация аутолизосом (Chen and Fang, 2002).Действительно, лечение рапамицином предотвращало индуцированную СКВ сывороточную активацию мембранных HLADR и CD86 (рис. S1), предполагая, что содержание сыворотки СКВ может увеличивать антигенпрезентирующую способность моноцитов посредством модуляции аутофагии.
и деградация аутолизосом (Chen and Fang, 2002).Действительно, лечение рапамицином предотвращало индуцированную СКВ сывороточную активацию мембранных HLADR и CD86 (рис. S1), предполагая, что содержание сыворотки СКВ может увеличивать антигенпрезентирующую способность моноцитов посредством модуляции аутофагии.
IFNα является важным цитокином, который, как было документально подтверждено, управляет DC-подобным фенотипом в моноцитах СКВ (Blanco et al., 2001). В соответствии с этим мРНК Cxcl10 (рис. S1), известный IFNα-индуцибельный ген, была активирована в моноцитах, обработанных сывороткой SLE, и степень их накопления SQSTM1 / P62 положительно коррелировала с концентрациями IFNα в сыворотке (рис. S1).Это побудило нас изучить, влияет ли передача сигналов IFN типа I напрямую на аутофагический поток здоровых моноцитов. Мы обнаружили, что аутофагия была индуцирована (A и 2B) уже через 4 часа, а накопление SQSTM1 / P62 последовало и достигло пика через 18 часов, достигая уровней накопления, наблюдаемых при лизосомном ингибировании гидроксихлорохином (CQ) (B – 2E и 2H). Лизосомная функция также была нарушена в моноцитах, обработанных IFNα, о чем свидетельствует значительно более низкая интенсивность LTR (F) и снижение активности катепсина D (G), в то время как обработка рапамицином восстанавливала лизосомную деградацию при передаче сигналов IFNα (F – 2H).
Лизосомная функция также была нарушена в моноцитах, обработанных IFNα, о чем свидетельствует значительно более низкая интенсивность LTR (F) и снижение активности катепсина D (G), в то время как обработка рапамицином восстанавливала лизосомную деградацию при передаче сигналов IFNα (F – 2H).
IFN-сигнал типа I ухудшает аутофаголизосомную деградацию в здоровых моноцитах CD14 +
CD14 + моноциты от здоровых доноров обрабатывали рекомбинантным IFNα (400 нг / мл) +/- CQ (46,1 мкΜ), рапамицином (рапамицином (рапамицином). ; 1 мкΜ), как показано.
(A) Относительная экспрессия мРНК ATG5 по сравнению с GAPDH (n = 9 здоровых доноров). Наборы данных были проанализированы с использованием непараметрического U-критерия Манна-Уитни.
(B) Вестерн-блоттинг на липидирование LC3B и уровни белка SQSTM1 / P62 в лизатах цельных клеток (n = 10 здоровых доноров).Показаны относительные интенсивности LC3II к LC3I и SQSTM1 / P62 к актину. Наборы данных были проанализированы с использованием непараметрического U-критерия Манна-Уитни.
(C) Относительное количество уровней мРНК SQSTM1 / P62, как в (A). Наборы данных были проанализированы с использованием непараметрического U-критерия Манна-Уитни.
(D) Репрезентативный вестерн-блот-анализ аутофагического потока при передаче сигналов IFNα.
(E) Конфокальная микроскопия для LC3B 555 , SQSTM1 / P62 488 , LAMP-1 633 и DAPI в моноцитах CD14 + от здоровых доноров, подвергшихся лечению, как указано.Изображен один репрезентативный результат. % совместной локализации LC3B и P62 puncta показан для моноцитов, культивированных в течение 18 часов (n = 8).
(F) Конфокальная микроскопия для Lysotracker DND99TM ± rapam. Изображен один репрезентативный результат. Показаны средние отношения интенсивностей флуоресценции (n = 6). Наборы данных были проанализированы с использованием парного критерия Стьюдента.
(G) Относительная активность катепсина D ± рапам. (n = 5). Наборы данных были проанализированы с использованием парного критерия Стьюдента.
(H) Анализ оценки аутофагического потока с помощью вестерн-блоттинга.Графически показано кратное изменение отношения интенсивностей P62 / актин (n = 4). Шкала 5 мкМ. Результаты выражены как среднее значение + стандартная ошибка среднего. p <0,05, p <0,005, p <0,0005. Наборы данных были проанализированы с использованием парного критерия Стьюдента.
Чтобы изучить функциональную важность IFNα-зависимых изменений лизосомной среды, которые нарушают завершение аутофагии в IFNα-DC, мы оценили их способность представлять антиген MHCII и секрецию провоспалительных цитокинов в присутствии рапамицина.По-видимому, добавление рапамицина снижает экспрессию мембран HLADR и CD86 и значительно снижает секрецию TNFα и интерлейкина (IL) 6 (A и 3B). Важно отметить, что обработанные IFNα моноциты CD14 + , подвергшиеся воздействию рапамицина, продемонстрировали заметно сниженную способность управлять наивной аллогенной пролиферацией CD4 + Т-клеток в реакции смешанных лимфоцитов (MLR), значительно улучшая их DC-подобный воспалительный фенотип (C). Чтобы выяснить, является ли этот эффект столь же значимым для иммунореактивности моноцитов СКВ, мы обработали их ex vivo рапамицином и выполнили MLR с наивными аллогенными CD4 + Т-клетками.Во всех случаях было обнаружено, что mTORC1 (мишень ингибирования рапамицина) активирован в свежевыделенных клетках от активных пациентов с СКВ по сравнению со здоровым контролем (D), а добавление рапамицина возвращало как их более высокие уровни HLADR и CD86, так и их способность праймировать T. клетки (E) более эффективно, чем здоровые моноциты (рисунок S2).
Чтобы выяснить, является ли этот эффект столь же значимым для иммунореактивности моноцитов СКВ, мы обработали их ex vivo рапамицином и выполнили MLR с наивными аллогенными CD4 + Т-клетками.Во всех случаях было обнаружено, что mTORC1 (мишень ингибирования рапамицина) активирован в свежевыделенных клетках от активных пациентов с СКВ по сравнению со здоровым контролем (D), а добавление рапамицина возвращало как их более высокие уровни HLADR и CD86, так и их способность праймировать T. клетки (E) более эффективно, чем здоровые моноциты (рисунок S2).
Нарушение аутолизосомальной деградации определяет повышенный иммуногенный потенциал IFNα-образных моноцитов и моноцитов СКВ.
CD14 + моноциты от здоровых доноров культивировали в течение 18 часов с IFNα (400 нг / мл) +/- рапам.(1 мкл), как показано.
(A) Уровни мембранной экспрессии HLA-DR и CD86 измеряли с помощью проточной цитометрии. Изображен репрезентативный результат. На графике нанесены средние геометрические значения интенсивности флуоресценции (GeoMFI) (n = 6).
(B) Концентрации секретируемых IL6 и TNFα, измеренные с помощью ELISA в супернатантах культур (n = 3).
(C) CFSE-меченные пуповинной крови CD4 + Т-клетки культивировали в течение 6 дней с аллогенными моноцитами, предварительно обработанными в течение 18 часов IFNα, +/- рапамом., и проанализированы на предмет их распространения. На графике представлены средние значения индекса пролиферации по 4 экспериментам. Справа: гистограммы разведения CFSE и размера (FS) закрытых клеток CD4 + . GeoMFI для непролиферирующих клеток и подсчет FS перечислены справа от гистограмм. Изображен один репрезентативный результат.
Экспрессия(D) p-4EBP1 и p-P70S6K в свежевыделенных моноцитах от здоровых (n = 5) и SLE (n = 5) доноров была проанализирована с помощью проточной цитометрии, и средние значения их GeoMFI представлены на графике.
(E) MLR между моноцитами СКВ ± рапам. (1 мкМ) в течение 1 часа, сокультивировали с аллогенными CFSE-меченными Т-клетками CD4 + , не имеющими отношения к пуповинной крови, в течение 6 дней (n = 3). Графически изображены средние значения индексов пролиферации, как в (C), и средние значения% HLADR hi CD86 hi популяции после сокультивирования.
Графически изображены средние значения индексов пролиферации, как в (C), и средние значения% HLADR hi CD86 hi популяции после сокультивирования.
(F) Типичная гистограмма интенсивности Lysosensor Green-189 при добавлении PLGA aNP в здоровые моноциты, культивируемые в течение 18 часов ± IFNα.
(G) Графики средних значений GeoMFI для HLADR и CD86 по данным проточной цитометрии (n = 3).Типичные гистограммы экспрессии HLADR и CD86 и диаграмма рассеяния, показывающая популяции HLADR hi CD86 hi .
(H) Показаны концентрации секретируемых IL6 и TNFα, измеренные с помощью ELISA в культуральных супернатантах тех же экспериментов. Результаты выражены как среднее значение + стандартная ошибка среднего. p <0,05, p <0,005. Все наборы данных были проанализированы с использованием парного критерия Стьюдента.
Известно, что помимо оптимизации лизосомного процессинга, модуляция активности mTORC1 оказывает плейотропное действие на дифференцировку ДК и презентацию антигена (Sukhbaatar et al. , 2016). Чтобы конкретно решить вопрос о том, что восстановление лизосомального pH может объяснить снижение IFNα-зависимой иммунореактивности моноцитов, мы использовали вместо них кислые наночастицы поли (DL-лактид- co -гликолид) (PLGA) (aNP) (Bourdenx et al., 2016). Эти биоразлагаемые полимеры, как известно, перемещаются в лизосомы и предотвращают подщелачивание лизосом даже в присутствии CQ. Действительно, совместная обработка здоровых моноцитов aNP и IFNα в течение 18 часов восстанавливала закисление лизосом (F) и значительно снижала мембранную экспрессию HLA-DR и CD86 (G) и секрецию IL6 и TNFα (H).
, 2016). Чтобы конкретно решить вопрос о том, что восстановление лизосомального pH может объяснить снижение IFNα-зависимой иммунореактивности моноцитов, мы использовали вместо них кислые наночастицы поли (DL-лактид- co -гликолид) (PLGA) (aNP) (Bourdenx et al., 2016). Эти биоразлагаемые полимеры, как известно, перемещаются в лизосомы и предотвращают подщелачивание лизосом даже в присутствии CQ. Действительно, совместная обработка здоровых моноцитов aNP и IFNα в течение 18 часов восстанавливала закисление лизосом (F) и значительно снижала мембранную экспрессию HLA-DR и CD86 (G) и секрецию IL6 и TNFα (H).
В совокупности метаболическое восстановление IFNα-опосредованного нарушения регуляции аутолизосом подавляет иммуногенный потенциал моноцитов, что указывает на важность эффективного аутофагического клиренса для улучшения иммунореактивности при передаче сигналов об опасности.
IFNα-опосредованная лизосомальная дисфункция препятствует клиренсу митохондрий и ведет к автономному накоплению мтДНК в клетках -ассоциированный молекулярный паттерн (DAMP), который может происходить из-за дефектной деградации автономного клеточного субстрата аутофагии.
 Важным внутренним интерферогенным DAMP, который физиологически перерабатывается посредством аутофагии, является мтДНК (Krysko et al., 2011). Таким образом, мы оценили митохондриальный гомеостаз и клиренс путем аутофагии (митофагии) в моноцитах СКВ. Окрашивание митотрекером (MTR) моноцитов активных, недавно диагностированных пациентов с СКВ показало повышенную среднюю интенсивность флуоресценции (MFI) и объем MTR + областей на клетку по сравнению с тем, что наблюдалось в здоровых моноцитах (A). Гиперполяризация митохондрий СКВ (увеличение потенциала митохондриальной мембраны [Δψ]) (Gergely et al., 2002a) может объяснить увеличение MTR MFI, но наблюдаемое увеличение объема MTR + областей на клетку повышает вероятность изменений в рециклинге митохондрий. Повреждение митохондрий распознается, на что указывает активация PINK1 (B), что индуцирует рекрутирование митохондрий в аутофагосомы. Чтобы проверить, правильно ли завершена митофагия, мы оценили количество мтДНК; мы количественно определили относительное количество копий мтДНК (область minArc), откалиброванных с помощью гена единственной копии геномной ДНК β2-микроглобулина (β2 m).
Важным внутренним интерферогенным DAMP, который физиологически перерабатывается посредством аутофагии, является мтДНК (Krysko et al., 2011). Таким образом, мы оценили митохондриальный гомеостаз и клиренс путем аутофагии (митофагии) в моноцитах СКВ. Окрашивание митотрекером (MTR) моноцитов активных, недавно диагностированных пациентов с СКВ показало повышенную среднюю интенсивность флуоресценции (MFI) и объем MTR + областей на клетку по сравнению с тем, что наблюдалось в здоровых моноцитах (A). Гиперполяризация митохондрий СКВ (увеличение потенциала митохондриальной мембраны [Δψ]) (Gergely et al., 2002a) может объяснить увеличение MTR MFI, но наблюдаемое увеличение объема MTR + областей на клетку повышает вероятность изменений в рециклинге митохондрий. Повреждение митохондрий распознается, на что указывает активация PINK1 (B), что индуцирует рекрутирование митохондрий в аутофагосомы. Чтобы проверить, правильно ли завершена митофагия, мы оценили количество мтДНК; мы количественно определили относительное количество копий мтДНК (область minArc), откалиброванных с помощью гена единственной копии геномной ДНК β2-микроглобулина (β2 m). Мы обнаружили накопление мтДНК в моноцитах СКВ по сравнению со здоровым контролем (В), что позволяет предположить либо усиление митохондриального биогенеза, либо ускользание мтДНК от митофагической деградации. Количественная оценка митохондриального белка VDAC1, который перерабатывается протеасомой как большинство белков внешней митохондриальной мембраны (Geisler et al., 2010), не выявила разницы между здоровыми моноцитами и моноцитами с СКВ (Рисунок S3). Это привело нас к возможности неэффективной деградации митохондриального содержимого, которое очищается митофагией.Примечательно, что передача сигналов IFNα индуцировала гиперполяризацию митохондрий, о чем свидетельствует увеличение внутримитохондриальных агрегатов красителя JC1 уже через 30 минут после обработки, и этот эффект сохранялся до 18 часов (D). Окрашивание MTR, экспрессия PINK1 и количественная оценка соотношений цикла количественной оценки (Cq) ядерной ДНК и митохондриальной ДНК дополнительно подтвердили, что передача сигналов IFNα повторяет фенотип моноцитов СКВ в отношении митохондриальной динамики и накопления мтДНК (E – 4G).
Мы обнаружили накопление мтДНК в моноцитах СКВ по сравнению со здоровым контролем (В), что позволяет предположить либо усиление митохондриального биогенеза, либо ускользание мтДНК от митофагической деградации. Количественная оценка митохондриального белка VDAC1, который перерабатывается протеасомой как большинство белков внешней митохондриальной мембраны (Geisler et al., 2010), не выявила разницы между здоровыми моноцитами и моноцитами с СКВ (Рисунок S3). Это привело нас к возможности неэффективной деградации митохондриального содержимого, которое очищается митофагией.Примечательно, что передача сигналов IFNα индуцировала гиперполяризацию митохондрий, о чем свидетельствует увеличение внутримитохондриальных агрегатов красителя JC1 уже через 30 минут после обработки, и этот эффект сохранялся до 18 часов (D). Окрашивание MTR, экспрессия PINK1 и количественная оценка соотношений цикла количественной оценки (Cq) ядерной ДНК и митохондриальной ДНК дополнительно подтвердили, что передача сигналов IFNα повторяет фенотип моноцитов СКВ в отношении митохондриальной динамики и накопления мтДНК (E – 4G). Добавление рапамицина предотвращало накопление мтДНК (G), тем самым демонстрируя, что IFNα-зависимые дефекты в разрешении митофагии препятствуют клиренсу митохондрий и приводят к автономному накоплению мтДНК в клетках.
Добавление рапамицина предотвращало накопление мтДНК (G), тем самым демонстрируя, что IFNα-зависимые дефекты в разрешении митофагии препятствуют клиренсу митохондрий и приводят к автономному накоплению мтДНК в клетках.Передача сигналов IFN типа I влияет на митохондриальный метаболизм, приводя к провоспалительному окислительному стрессу — mtROS и АТФ влияют на лизосомный pH
Чтобы проанализировать молекулярный механизм, регулирующий аберрантность СКВ при деградации лизосом, мы выполнили секвенирование следующего поколения на основе моноцикла (RNA-setesqto) активные, нелеченые пациенты с СКВ (таблицы S3 и S4). Анализ пути показал, что сигнатура IFN типа I (репрезентативные гены IFI27, USP18, IFITM1, MX1), увеличение презентации и дифференцировки антигена (MMP10, IL6, CSF3, MIR155HG, SIGLEC-1) совпадают с ответом на окислительный стресс и изменениями функции митохондрий. и метаболизм (ACOT4, CYP19A1, MOXD1, IFI6, GMPR), а также в регуляции рН эндоцитов и ионных каналов (IGLL5, CLIC5, ATP10A, KCNh4, CACNB4) (A; Таблица S1).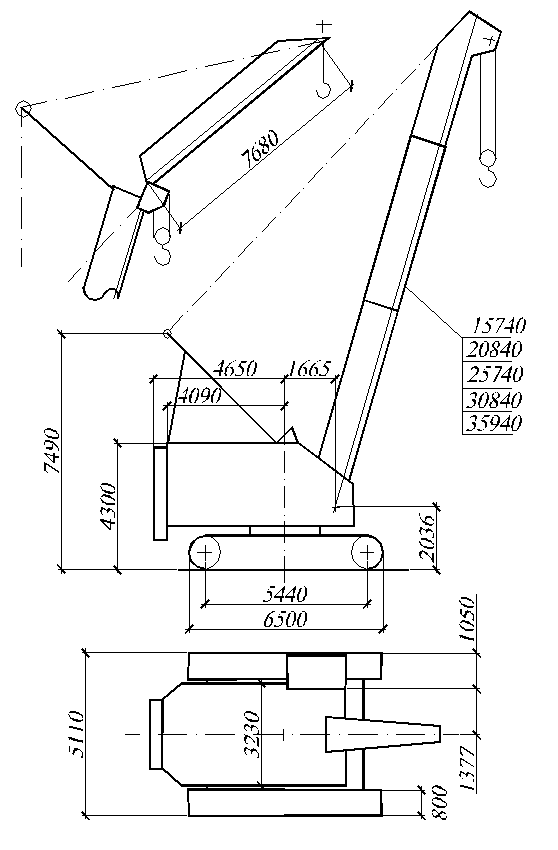 Эти результаты побудили нас сформулировать гипотезу о том, что IFNα-опосредованная дерегуляция митохондриального метаболизма влияет на pH лизосом, что приводит к нарушению клиренса митохондрий. На стабильность лизосомного pH влияет локализация и активность субъединицы mTORC1, которая функционирует, чтобы связать клеточное метаболическое состояние с лизосомной деградацией посредством восприятия АТФ (Cang et al., 2013). После подтверждения того, что mTORC1 активируется при лечении IFNα (Matsumoto et al., 2009), мы выяснили, может ли аберрантный OXPHOS, о чем свидетельствует стойкая гиперполяризация митохондрий, сочетаться с IFNα-зависимым подщелачиванием лизосом через определение АТФ.Мы оценили уровни как АТФ, так и mtROS в здоровых моноцитах, культивированных с IFNα. Уровни mtROS были увеличены уже через 2 часа после обработки IFNα, тогда как уровни АТФ были снижены, и этот эффект сохранялся до 18 часов, что подтверждается окрашиванием MitoSOX и колориметрической оценкой внутриклеточной концентрации АТФ (B и 5C).
Эти результаты побудили нас сформулировать гипотезу о том, что IFNα-опосредованная дерегуляция митохондриального метаболизма влияет на pH лизосом, что приводит к нарушению клиренса митохондрий. На стабильность лизосомного pH влияет локализация и активность субъединицы mTORC1, которая функционирует, чтобы связать клеточное метаболическое состояние с лизосомной деградацией посредством восприятия АТФ (Cang et al., 2013). После подтверждения того, что mTORC1 активируется при лечении IFNα (Matsumoto et al., 2009), мы выяснили, может ли аберрантный OXPHOS, о чем свидетельствует стойкая гиперполяризация митохондрий, сочетаться с IFNα-зависимым подщелачиванием лизосом через определение АТФ.Мы оценили уровни как АТФ, так и mtROS в здоровых моноцитах, культивированных с IFNα. Уровни mtROS были увеличены уже через 2 часа после обработки IFNα, тогда как уровни АТФ были снижены, и этот эффект сохранялся до 18 часов, что подтверждается окрашиванием MitoSOX и колориметрической оценкой внутриклеточной концентрации АТФ (B и 5C). Удаление mtROS с помощью MitoTEMPO предотвращало опосредованное IFNα снижение образования АТФ и дополнительно увеличивало его продукцию по сравнению с базовыми уровнями (C) и восстанавливало лизосомный pH (D).Добавление рапамицина вернуло нарушенное равновесие продукта OXPHOS, восстановив продукцию mtROS до физиологических уровней, а также повысив уровни АТФ (B и 5C). В подтверждение, скорость потребления кислорода, которая была значительно ниже при передаче сигнала IFNα, была восстановлена добавлением рапамицина или улавливания mtROS (фигура S6). В совокупности наши результаты предполагают существование порочного круга дефектного митохондриального дыхания и окислительного стресса, который генерируется и усиливается при передаче сигналов IFNα.Гиперполяризация митохондрий, указывающая на дефектный OXPHOS, увеличивает соотношение mtROS-ATP и это ухудшает лизосомную деградацию, что, в свою очередь, препятствует эффективному клиренсу поврежденных митохондрий, тем самым подавляя способность моноцитов контролировать окислительный стресс.
Удаление mtROS с помощью MitoTEMPO предотвращало опосредованное IFNα снижение образования АТФ и дополнительно увеличивало его продукцию по сравнению с базовыми уровнями (C) и восстанавливало лизосомный pH (D).Добавление рапамицина вернуло нарушенное равновесие продукта OXPHOS, восстановив продукцию mtROS до физиологических уровней, а также повысив уровни АТФ (B и 5C). В подтверждение, скорость потребления кислорода, которая была значительно ниже при передаче сигнала IFNα, была восстановлена добавлением рапамицина или улавливания mtROS (фигура S6). В совокупности наши результаты предполагают существование порочного круга дефектного митохондриального дыхания и окислительного стресса, который генерируется и усиливается при передаче сигналов IFNα.Гиперполяризация митохондрий, указывающая на дефектный OXPHOS, увеличивает соотношение mtROS-ATP и это ухудшает лизосомную деградацию, что, в свою очередь, препятствует эффективному клиренсу поврежденных митохондрий, тем самым подавляя способность моноцитов контролировать окислительный стресс.
Передача сигналов IFN типа I влияет на митохондриальный метаболизм, приводя к провоспалительному окислительному стрессу и ионным изменениям лизосомных компартментов 7) и здоровый контроль (n = 5) CD14 + моноциты.Все недифференциально экспрессируемые гены показаны серым цветом. Выбранные гены, относящиеся к сигнатуре IFN I типа, окислительному стрессу, метаболизму, презентации антигена и дифференцировке, отмечены в верхнем (красный цвет) и вниз (синий цвет) генах, по-разному экспрессируемых.
(B) Относительный митохондриальный O 2 — уровни CD14 в здоровых моноцитах + ± IFNα, рапам. были оценены с помощью проточной цитометрии с использованием красителя mitoSOX (n = 5). Показано наложение одной репрезентативной гистограммы и показаны ворота, указывающие на низкий и высокий уровни mtROS.Графически представлены средние значения% клеток в популяции mitoSOX high. Наборы данных были проанализированы с использованием парного критерия Стьюдента.
(C) Концентрация внутриклеточной концентрации АТФ ± IFNα, rapam. Или mitoTEMPO в лизатах моноцитов CD14 + (n = 5). Наборы данных были проанализированы с использованием парного критерия Стьюдента.
(D) Конфокальная микроскопия для Lysotracker DND99 в моноцитах CD14 + от здоровых доноров ± IFNα, mitoTEMPO. Изображен один репрезентативный результат. Графически представлены средние значения MFI на ячейку (n = 3).Наборы данных были проанализированы с использованием парного критерия Стьюдента.
(E) Уровни экспрессии HLA-DR и CD86, измеренные с помощью проточной цитометрии в моноцитах CD14 + , при передаче сигнала IFNα и улавливании mtROS. Построены средние значения GeoMFI (n = 3). Наборы данных были проанализированы с использованием парного критерия Стьюдента.
(F) Концентрации секретируемых IL6 и TNFα, измеренные с помощью ELISA, в культуральных супернатантах моноцитов CD14 + (n = 7), культивируемых, как в (D), представлены на графике. Шкала 5 мкМ. Результаты выражены как среднее значение + стандартная ошибка среднего. p <0,05, p <0,005.
Шкала 5 мкМ. Результаты выражены как среднее значение + стандартная ошибка среднего. p <0,05, p <0,005.
Чтобы выяснить, влияют ли IFNα-опосредованные эффекты на митохондриальное дыхание на воспалительное прайминг моноцитов при передаче сигналов IFNα, мы собрали mtROS и оценили их антигенпредставляющие свойства. Добавление MitoTEMPO предотвращало IFNα-зависимое увеличение мембранной экспрессии HLA-DR и, в меньшей степени, CD86 (E) и сопутствующей секреции IL6 и TNFα (F), тем самым улучшая эффекты провоспалительного внутриклеточного окислительного стресса.
Аутофагия ускользания мтДНК ощущается через STING и запускает моноциты для аутоиммунного ответа
Аутофагическое ускользание мтДНК является клеточно-автономным триггером, который задействует противовирусную передачу сигналов для усиления экспрессии подмножества стимулированных IFN генов и может вызывать воспаление и патологию (Ока и др., 2012). Неметилированная собственная ДНК, цитоплазматическое высвобождение которой не связано с запрограммированной гибелью клеток, может быть обнаружена TLR9 в эндосомах и цитозольными сенсорами (DAI, IFI16 / P202, DDX41 и cGAS) (de Jong et al. , 2010, Klinman et al., 1996, West et al., 2015), которые активируют адаптерную молекулу STING. Как показали наши данные РНК-seq на моноцитах СКВ активных пациентов с сильной сигнатурой IFNα, цитозольные сенсоры ДНК сверхэкспрессируются (DAI, IFI16, ZBP1), тогда как cGAS, который принадлежит к семейству IFN-стимулированных генов (ISG) (Ma et al. al., 2015), дифференцированно не выражалась. Чтобы определить, какой путь задействован во время восприятия мтДНК при передаче сигналов IFNα, и перевести его в повышенную иммуногенность, мы подавили экспрессию TLR9 и STING в здоровых моноцитах, культивировали с GMCSF и IL4 или IFNα в течение 3 дней, и измерили их антигенпрезентирующую способность.Мы обнаружили, что IFNα-опосредованное увеличение моноцитов HLA — DR + CD86 + и секреция CXCL10 были значительно снижены при нокдауне STING, тогда как при молчании TLR9 не было обнаружено значительных различий (A и 6B). Этот эффект был специфическим для IFNα, поскольку нокдаун STING не влиял на экспрессию HLA-DR и CD86 или секрецию CXCL10 моноцитами, обработанными IL-4.
, 2010, Klinman et al., 1996, West et al., 2015), которые активируют адаптерную молекулу STING. Как показали наши данные РНК-seq на моноцитах СКВ активных пациентов с сильной сигнатурой IFNα, цитозольные сенсоры ДНК сверхэкспрессируются (DAI, IFI16, ZBP1), тогда как cGAS, который принадлежит к семейству IFN-стимулированных генов (ISG) (Ma et al. al., 2015), дифференцированно не выражалась. Чтобы определить, какой путь задействован во время восприятия мтДНК при передаче сигналов IFNα, и перевести его в повышенную иммуногенность, мы подавили экспрессию TLR9 и STING в здоровых моноцитах, культивировали с GMCSF и IL4 или IFNα в течение 3 дней, и измерили их антигенпрезентирующую способность.Мы обнаружили, что IFNα-опосредованное увеличение моноцитов HLA — DR + CD86 + и секреция CXCL10 были значительно снижены при нокдауне STING, тогда как при молчании TLR9 не было обнаружено значительных различий (A и 6B). Этот эффект был специфическим для IFNα, поскольку нокдаун STING не влиял на экспрессию HLA-DR и CD86 или секрецию CXCL10 моноцитами, обработанными IL-4.
Аутофагический выход мтДНК ощущается с помощью STING и инициализирует моноциты для аутоиммунного ответа
(A) Моноциты CD14 + от здоровых доноров (n = 5) подвергали электропорации с помощью скремблирования, TLR9 и STING малых интерферирующих РНК (миРНК) и культивировали в течение 3 дней с GMCSF и IL4 или IFNα.Уровни экспрессии HLA-DR и CD86 анализировали с помощью проточной цитометрии. На графике представлены средние значения% популяций HLADR hi CD86 hi .
(B) Секреция CXCL10 как совокупный индикатор передачи сигналов IFNα при скремблировании, STING и нокдауне TLR9.
(C) Секреция IL6 и TNFα в супернатантах культур (n = 3).
(D) Относительные уровни мРНК STING по сравнению с GAPDH в свежевыделенных моноцитах CD14 + от (n = 5) здоровых доноров и (n = 6) пациентов с активной СКВ.Результаты выражены как среднее значение + стандартная ошибка среднего. p <0,005.
(E) Моноциты CD14 + от здоровых доноров (n = 4), трансфицированные функциональным или инактивированным нагреванием белком ДНКазы I ± IFNα. Уровни экспрессии HLA-DR и CD86 измеряли с помощью проточной цитометрии. Изображены типичные гистограммы количества HLADR и CD86 между клетками, обработанными IFNα-ДНКазой I, и клетками, обработанными термоинактивированной ДНКазой I. Графически представлены средние значения GeoMFI. Результаты выражены как среднее значение + стандартная ошибка среднего. р <0.05, p <0,005. Наборы данных были проанализированы с использованием парного критерия Стьюдента, кроме (D), который был проанализирован с использованием непараметрического U-критерия Манна-Уитни.
Интересно, что нокдаун STING существенно снизил опосредованную IFNα секрецию IL6, в то время как секреция TNFα также снизилась, но не достигла статистической значимости (p = 0,0626) (C), что свидетельствует о том, что зондирование STING может контролировать опосредованный интерфероном I типа ответ на иммуногенную нуклеиновую кислоту. наличие в цитоплазме. Эти результаты согласуются с повышенной экспрессией STING, наблюдаемой в моноцитах СКВ (D), и повторяют эффект ингибирования mTORC1, повторного подкисления лизосом и поглощения mtROS при передаче сигналов IFNα.
Наконец, чтобы подтвердить, что цитозольный выход мтДНК из автолизосом ощущается с помощью STING при передаче сигналов IFNα, мы трансфицировали моноциты независимым от кислотности белком ДНКазой I, используя реагент для трансфекции DOTAP, липоплекс, который интернализуется клатрин-опосредованным путем эндоцитоза и направлен в лизосомный компартмент (Rejman et al., 2005). Трансфекция ДНКазой I снижала аутовоспалительное IFNα-опосредованное увеличение антигенпредставляющей способности моноцитов на основе экспрессии HLADR и CD86 по сравнению с доставкой инактивированной нагреванием ДНКазы I (E).Эти данные подтверждают, что неэффективная деградация субстрата лизосомальной нуклеиновой кислоты ощущается и стимулирует ответ.
Обсуждение
Рациональная разработка альтернативных таргетных методов лечения системного аутоиммунного заболевания требует всестороннего понимания патогенных молекулярных механизмов, обращение которых может привести к восстановлению гомеостаза. Неправильная очистка и накопление биологических отходов вместе с восприятием собственной ДНК и чрезмерным продуцированием IFNα стимулируют патогенез СКВ (Crow, 2014, Niewold et al., 2007, Паскуаль и др., 2006, Цокос, 2016). В этом контексте было получено дополнительное понимание нашего понимания путей и сенсоров врожденного иммунитета, которые отвечают за инициирование аутовоспалительных событий, запускаемых собственной ДНК, в иммунных клетках (Caielli et al., 2016, Oka et al., 2012, Shrivastav and Niewold, 2013). Тем не менее, остается неясным, может ли и как устойчивая передача сигналов IFNα влиять на непрерывную продукцию этих DAMP собственной ДНК для сохранения аутоиммунного ответа, регулирующего СКВ.Наша работа предполагает критическую роль IFNα-опосредованного накопления мтДНК в моноцитах СКВ, которая избегает аутофагического клиренса и действует как клеточно-автономный цитоплазматический DAMP. Его восприятие и активация STING приводит к усилению экспрессии MHCII и CD86 и секреции TNFα и IL6, что позволяет им запускать иммунитет к собственной ДНК и способствовать клональной экспансии аутореактивных лимфоцитов.
Наши результаты продемонстрировали, что IFNα индуцирует аутофагию в наивных моноцитах CD14 + , наблюдение согласуется с сообщениями об индукции аутофагии IFN типа I в нескольких линиях раковых клеток (Schmeisser et al., 2013, Zhu et al., 2015). Анализ промотора LC3B человека in silico показал наличие участков связывания Stat и IRF, и было обнаружено, что транскрипты гена LC3B активированы в обработанных IFNα моноцитах (рис. S4), что указывает на возможность прямой регуляции транскрипции компонентов аутофагии с помощью IFNα. Помимо индукции аутофагии, IFNα нарушает завершение аутофагического потока, моноцитарные лизосомы не могут должным образом подкисляться, что приводит к снижению переваривания лизосомных субстратов и накоплению поврежденных субстратов аутофагии в воспринимаемой цитоплазме.Анализ уровней экспрессии TFEB и ядерной локализации, фактора транскрипции, который контролирует лизосомно-аутофагический путь (Settembre et al., 2011) и управляет биогенезом лизосом, вместе с количественной оценкой точки LAMP1 на клетку не показал различий при СКВ и при лечении IFNα (Рисунок S5), что еще раз подтверждает, что дефицит аутофагической деградации является результатом нарушения регуляции лизосомной активности.
Наши данные анализируют метаболический путь, запускаемый IFNα, который приводит к замедлению лизосомного переваривания за счет изменения его регуляции pH.В соответствии с предыдущим исследованием с использованием СКВ PBMC (Gergely et al., 2002b) мы наблюдали, что моноциты СКВ несут более низкие уровни АТФ и более высокие уровни mtROS, что свидетельствует о высоком митохондриальном мембранном потенциале, который является результатом дерегуляции OXPHOS (Рисунок S6). Наши данные РНК-секвенирования моноцитов СКВ с высокой сигнатурой IFNα указывают на нарушение митохондриального метаболизма, окислительный стресс и изменения в ионных каналах, а также в регуляции эндосомального pH. В состоянии митохондриальной гиперполяризации, как это видно в СКВ PBMC, ионы H + вытесняются из митохондриального матрикса в цепи переноса электронов (ETC) — и цитохромы восстанавливаются, способствуя выработке ROS и вызывая окислительный стресс, как указано. за счет активации генов AXL, FASLG, HERC5, ABTB2, ETS1, FBXO2, HERC6, IL1A, PARP15, PARP9, SPSB1 и TRIM69 в моноцитах СКВ.
Иммунометаболизм — это новый рубеж в иммунологии (Mathis and Shoelson, 2011). Известно, что выживание и гиперактивация Т-клеток СКВ поддерживается интенсивным OXPHOS (Wahl et al., 2012). Нормализация метаболизма CD4 + Т-клеток у мышей с предрасположенностью к волчанке была предложена в качестве многообещающей стратегии обращения вспять патологии волчанки (Yin et al., 2015). OXPHOS и его продукты, АТФ и mtROS, являются основными детерминантами внутриклеточного окисления и регулируют клеточную активацию, дифференциацию и выживание.Те же побочные продукты митохондриального дыхания модулируют аутофагию и ее поток. mTOR представляет собой ΔΨm-сенсор митохондриальной мембраны, который может активироваться окислительно-восстановительным или АТФ-зависимым образом, и его блокада стала эффективным подходом к обратной активации Т-клеток (Pearce et al., 2013, Sena et al., 2013, Yin и др., 2015). Находясь на лизосомной мембране, субъединица mTORC1 mTOR обладает способностью регулировать лизосомный pH и определять его функциональность (Korolchuk et al., 2011). Метаболическая регуляция функции лизосом влияет на разрешение аутофагической деградации, влияя на судьбу его субстратов, модулируя активацию и дифференцировку иммунных клеток (Chen et al., 2008). Наше исследование обеспечивает ранее не охарактеризованный механизм и раскрывает компоненты, влияющие на иммунометаболизм моноцитов при СКВ человека, которые могут быть использованы в качестве дополнительной терапии для ускорения ремиссии и предотвращения обострений заболевания. Кислотные наночастицы используются в качестве инструментов для коррекции эффектов аберрантного иммунометаболизма, становясь более селективным вариантом лечения СКВ. В частности, aNP PLGA также могут разрушаться, их производные (молочная и гликолевая кислоты) могут поддерживать Δψ mt., и их действие остается неизменным в присутствии гидроксихлорохина, основного препарата для лечения волчанки.
Метаболическое восстановление IFNα-зависимой дерегуляции закисления лизосом приводит к завершению аутофагии и снижает иммуногенный потенциал IFNα-индуцированных моноцитов. Передача сигналов IFΝα напрямую влияет на функцию митохондрий и OXPHOS, на что указывает повышенное присутствие Stat1 в митохондриях моноцитов, обработанных IFNα (фиг. S6), которое, как ранее сообщалось, влияет на Δψ m .это объясняет — по крайней мере частично — наблюдаемую гиперполяризацию митохондрий СКВ с последующим повышением генерации mtROS и снижением АТФ. Окислительный стресс ощущается, на что указывает активация гена PINK1, отвечающего на АФК, индуцируется аутофагия, но лизосомная дисфункция, опосредованная IFNα, препятствует клиренсу митохондрий, что приводит к автономному накоплению мтДНК в клетках, которая активирует STING и подготавливает моноциты для защиты от неправильно локализованных бактериоподобных ядер. кислоты. Наблюдаемое функциональное восстановление деградации лизосом и связанное с этим улучшение их воспалительного фенотипа позволяет предположить, что ингибирование mTORC1 может обратить воспалительную функцию IFNα-образных моноцитов.Аналогичные эффекты были достигнуты путем воздействия на любой этап предложенного механизма, такого как mtROS через поглощение, лизосомный pH за счет подкисления с помощью PLGA aNP, расщепление ДНК посредством трансфекции ДНКазой I в лизосомы или нокдаун STING в IFNα-образных моноцитах, тем самым препятствуя опосредованному IFNα дифференцировка моноцитов крови в DC-подобные APC.
Ингибиторы mTOR, такие как сиролимус и эверолимус, эффективны как часть схем тройной иммуносупрессии для предотвращения отторжения трансплантата почки, причем их основными преимуществами являются отсутствие нефротоксичности и снижение частоты злокачественных новообразований.Использование ингибиторов mTOR при СКВ (как у человека, так и у мышей) снизило тяжесть нефрита и общую активность заболевания, а также продукцию антифосфолипидных антител и сосудистые поражения, связанные с антифосфолипидным синдромом (Bride et al., 2016, Canaud et al., 2014, Oaks et al., 2016). Последние данные многоцентрового исследования показывают, что рапамицин эффективен при рецидивирующей / рефрактерной аутоиммунной цитопении (Chen and Fang, 2002). Несмотря на эти обнадеживающие результаты, необходимо лучше задокументировать роль ингибирования mTOR в лечении СКВ.На сегодняшний день их положительные эффекты приписываются их отменяющему действию на активацию mTOR в эндотелиальных клетках почечных сосудов и нормализации индуцированного активацией Т-клеток потока кальция (Crow and Manel, 2015). Здесь мы демонстрируем дополнительный эффект на лизосомный клиренс в моноцитах СКВ — критическое событие как при возникновении, так и при рецидиве заболевания.
Наше исследование установило, что гиперполяризация митохондрий, наблюдаемая в моноцитах СКВ, может быть опосредована стимуляцией IFNα, которая активирует механизм, связывающий передачу сигналов цитокинов с OXPHOS.Обращение IFα-зависимого внутриклеточного окисления с использованием поглотителя mtROS представляет собой дополнительную цель, которую необходимо оценить с точки зрения его эффективности в достижении стойкой ремиссии — основной цели в лечении СКВ. Антиоксидантные поглотители mtROS в настоящее время исследуются при нескольких заболеваниях, связанных с митохондриальной дисфункцией и окислительным стрессом, таких как антифосфолипидный синдром, рассеянный склероз и васкулит. Их эффективность в поддержании гомеостаза нейрональных и эндотелиальных клеток, в то время как они одновременно действуют как мягкие иммунодепрессанты, делает их привлекательными кандидатами для предотвращения или улучшения патологии СКВ, включая почечные, психоневрологические и скелетно-мышечные проявления.
Мульти-направленная терапия необходима для достижения ремиссии и предотвращения обострений СКВ. Наши данные предполагают неожиданные преимущества нацеливания на IFNα при СКВ и выявляют дополнительные терапевтические мишени, которые нарушают «порочный» цикл аберрантного восприятия нуклеиновых кислот в моноцитах волчанки и, таким образом, могут способствовать более глубокой и длительной ремиссии при этом заболевании. Разработка сложных инструментов и подходов к in vivo регулирует молекулярные аберрации, которые вызывают патологию, может открыть путь для профилактики и методического лечения сложных заболеваний человека.
STARMethods
Таблица основных ресурсов
| РЕАГЕНТ или РЕСУРС | ИСТОЧНИК | ИДЕНТИФИКАТОР | |||
|---|---|---|---|---|---|
| Кроличьи 9088 | Каталожный № NB100-2220; RRID: AB_10003146 | ||||
| Мышиные моноклональные анти-LC3B (клон 5F10) | Nanotools | Кат. № 0231-100; RRID: AB_2722733 | |||
| Кролик поликлональный анти-SQSTM1 / P62 | MBL | Кат. № PM045; RRID: AB_1279301 | |||
| Кроличьи поликлональные анти-VDAC | Millipore | Кат. № AB10527; RRID: AB_10806766 | |||
| Крысиный монокломальный анти-LAMP1 (клон 1D4B) | Abcam | Каталожный номер ab25245; RRID: AB_449893 | |||
| анти-HLADR (клон L243) | Biolegend | Кат. № 30762728 | |||
| анти-CD86 (клон IT2.2) | Biolegend | Cat # 30540506 | |||
| anti-CD80 (clone 2D10) | Biolegend | Cat # 305219 | |||
| anti-CD14 (clone HCD14) | Biolegend | Biolegend | |||
| анти-pS6K PECy7 (клон CUPK43K) | eBioscience | Кат. № 25 1 | |||
| анти-p4EBP1 PE (клон V3NTY24) | invitrogen | -99097Химические вещества, пептиды и рекомбинантные белки | |||
| Универсальный интерферон I типа, человеческий | PBL | Кат. № 11200-1 | |||
| Pepbinant6 Pepbinant6, человеческий IL-4 | # 200-04 | ||||
| Рекомбинантный GMCSF человека | Peprotech | Кат. № 300-03 | |||
| Резомер RG 503 H, поли (D, L-лактид-со-глик) олид) с концевыми кислотными группами, лактид: гликолид 50:50, Mw 24,000-38,000 | SIGMA | Номер по каталогу: 719870 | |||
| Критические коммерческие анализы | Abcam | Кат. № ab65302 | |||
| Набор для анализа АТФ | Abcam | Кат. | |||
| Набор для выделения митохондрий для культивируемых клеток | Thermo Scientific | Кат. № 89874 | |||
| VeriKine Human Interferon Alpha Multi-Subtype Serum ELISA Kit | PBL | Human | Cat # 41110-1 909 Set -Go ELISA | eBioscience | Cat # 887346 |
| Human IL6 Ready-Set-Go ELISA | eBioscience | Кат. № 887066 | |||
| Человеческий CXCL10 (IP-10) ИФА | eBioscience | Кат. | |||
| Депонированные данные | |||||
| Данные РНК-seq, SRA Bioproject PRJNA392602 | Краткий архив чтения NCBI | https: // www.ncbi.nlm.nih.gov/bioproject/PRJNA392602/ | |||
| Олигонуклеотиды | |||||
| TM Human ssteps7 HSS139158 | Life Technologies | Cat # 1299003 | |||
| Stealth siRNAs (набор из 3), TLR9 Human, HSS147622, HSS147623, HSS147624 | Life Technologies | Cat # 12990038 siRNA | Life TechnologiesКат. № 12935-100 | ||
| Праймеры для IFNα, CXCL10, SQSTM1 / P62, LC3, PINK1, Parkin, ATG5, huβ2Μ, mtMinArc (методы STAR) | эта статья | 6 N /Праймеры и зонд STING (TMEM173) (FAM): Hs00736958_m1 assay | Life Technologies | Каталожный номер 4331182 | |
| Праймеры и зонд GAPDH (HEX) Hs03929097_g1 | Li fe Technologies | Кат. № 4448489 | |||
| Праймеры и зонд TLR9: Hs00152973_m1 (FAM) | Life Technologies | Кат. | |||
| Призма GraphPad | НЕТ | https: // www.graphpad.com | |||
| Fiji | https://fiji.sc/ | ||||
| Макрос для% колокализации для программного обеспечения Fiji | Daniel J. Shiwarski | N / A | |||
Другое | |||||
| CD14 MicroBeads, человек | Miltenyi | Кат. -091-153 | |||
| CD4 + CD25 + Набор для выделения регуляторных Т-лимфоцитов, человек | Miltenyi | Кат. № 130-091-301 | |||
| Лизосенсор зеленый DND-189 | Life Technologies | LysoTracker Red DND-99 | Life Technologies | Cat # L7528 | |
| Mitotracker Red | Life Technologies | Cat # M7512 | |||
| Набор для пролиферации клеток CellTrace CFSE | Life Technologies | Кат. № C34554 | |||
| Красный индикатор митохондриального супероксида MitoSOX | Life Technologies | Life Technologies | Кат. | ||
| Рапамицин | SIGMA | Каталожный номер 553210 EMD Millipore | |||
| Плаквенил (гидроксихлорохин) | Sanofi Aventis | Код ATC 8P01BA02 | липосагент 911BA02 | липос 9999 9998 9989 | липосКонтактное лицо для обмена реагентами и ресурсамиДополнительная информация и запросы на ресурсы и реагенты должны быть направлены и будут выполнены ведущим контактным лицом, Катериной Гкиртзиманаки (kgirtzimanaki @ gmail.com). Экспериментальная модель и детали предметаЛюдиБыли изучены 49 пациентов с СКВ, которые находились в ревматологической клинике Университетской больницы Крита, диагноз поставлен в соответствии с критериями классификации Американского колледжа ревматологии 1982 года (средний возраст = 42,5 года; диапазон: От 16 до 70 лет). Активная СКВ определялась как показатель индекса активности СКВ 2000 (SLEDAI-2K) выше 8 в день забора крови. Пациенты воздерживались от приема лекарств, включая стероиды и CQ, по крайней мере за 24 часа до забора крови.Более того, n = 26 пациентов полностью прекратили терапию на момент включения в исследование. Добровольцы того же возраста и пола здорового возраста из отделения трансфузиологии университетской больницы Ираклиона служили контролем. 6 пациентов, прошедших лечение в ревматологической клинике Университетской клиники Крита, с диагнозом ревматоидный артрит в соответствии с критериями Американского колледжа ревматологии (ACR) 1987 г., сдали периферическую кровь, которая использовалась в качестве контрольной сыворотки (средний возраст = 45 лет).4 года). Исследование было одобрено этическим комитетом университетской больницы Ираклиона, и все субъекты дали письменное информированное согласие перед исследованием (номер протокола: 5944 / 14-6-2017). Пол / пол, возраст и истории болезни пациентов с СКВ перечислены в таблице S1 (файл Excel), а пациентов с РА — в таблице S2. Критериями отбора пациентов были активность заболевания (SLEDAI) и способность воздерживаться от терапии. Подробные сведения о методахВыделение моноцитов периферической кровиPBMC выделяли из гепаринизированной периферической крови человека с помощью центрифугирования с плотностью фиколл-Histopaque-1077 (GE-Healthcare).Эритроциты удаляли с помощью гипотонического лизиса (1 мл ddh3O в течение 25 с и 1 мл 1,8% NaCl). Жизнеспособность была измерена на 99% по исключению красителя трипанового синего в каждом эксперименте. Моноциты выделяли с помощью магнитного поля (MACS, Miltenyi Biotec) и чистоту оценивали с помощью проточной цитометрии (CD14 + клетки / все живые клетки). Во всех экспериментах использовали препараты чистотой ≥ 94%. Сбор сывороткиОбразцы периферической крови здоровых людей, больных СКВ и РА собирали в вакуумеры SST и центрифугировали при 2500 об / мин в течение 15 минут (RT).Сыворотку собирали в стерильных условиях и хранили в аликвотах при -80 ° C. Клеточная культураМагнитно изолированные моноциты CD14 + культивировали в RPMI-1640 / L-глутамине с добавлением 10% термоинактивированного FBS, 100 МЕ / мл пенициллина и 100 мкг / мл стрептомицина в течение 24 часов. Когда требовалось более длительное время культивирования, клетки культивировали в течение 3 дней, а в среду дополнительно добавляли GM-CSF (100 нг / мл, Peprotech) и IL-4 (20 нг / мл, Peprotech) или универсальный IFN типа I (Hu- IFN-αA / D [Bg / II] — 400 нг / мл, PBL) или гетерологичная здоровая сыворотка, SLE или RA (10% об. / Об. В бессывороточной среде). Проточная цитометрияЖивые клетки (набор для апоптоза аннексина V, BD PharMingen) окрашивали антителами, конъюгированными с флуорохромом (клон HCD14 против CD14, клон Okt4 против CD4, клон IT2.2 против CD86, клон L243 против HLADR, клон T36 / 45 против p4EBP1, клон против pP70S6K Cl 215247) в течение 20 мин при 4 ° C в PBS / 5% FBS. АФК в митохондриях измеряли с помощью окрашивания MitoSOX (Invitrogen) (1 мМ в течение 15 минут при 37 ° C и 5% CO 2 ). Поляризацию митохондриальной мембраны оценивали с помощью окрашивания JC-1 (eBiosciences) (1 мкг / мл в течение 10 минут при 37 ° C и 5% CO2), а повторное закисление лизосом с помощью aNP оценивали с помощью.Окрашивание лизосенсором DND189 (Thermo Scientific) (1 мкл в среде в течение 20 минут при 37 ° C и 5% CO2). Образцы собирали на FACSCalibur (BD Biosciences) и анализировали с помощью программного обеспечения FlowJo (Tree Star). Для окрашивания фосфобелков клетки пермеабилизировали и окрашивали с использованием набора буфера для внутриклеточной фиксации и пермеабилизации (eBioscience) в соответствии с инструкциями производителя. Реакция смешанных лимфоцитовПуповинная кровь человека (CB) была получена при нормальных родах акушерско-гинекологическим отделением университетской больницы Ираклиона.CB собирали в гепаринизированную пробирку после родов, пережатия и пересечения пупка. CD4 + CD25 — Т-клетки были изолированы с помощью магнитного поля (Miltenyi, CD4 + CD25 + набор для выделения Treg 130-091-301, отрицательная селекция) с помощью CB PBMC (чистота ≥ 85%) и маркированных CFSE (Thermo Fisher Scientific, Cell Trace C34554). Аллогенные моноциты (Miltenyi, набор для выделения моноцитов II, 130-091-153, отрицательный отбор) были изолированы от неродственных взрослых доноров PBMC (чистота ≥ 94%) за 1 день до донорства CB в случае здоровых доноров или в тот же день в случае случаи больных СКВ; Типирование HLA не производилось.Здоровые CD14 + PBMC предварительно обрабатывали в течение 18 часов следующими реагентами, по отдельности или в комбинации: универсальный IFN типа I (IFNα), рапамицин и CQ, и промывали перед добавлением в MLR. Моноциты СКВ предварительно обрабатывали рапамицином в течение 1 часа, если это указано. 10 5 предварительно обработанных моноцитов помещали в 96-луночные планшеты с круглым дном и добавляли 5×10 4 Т-клеток, меченных CFSE; MLR оставляли в культуре на 6 дней в увлажненном инкубаторе с температурой 37 ° C и 5% CO 2 .Каждую культуру MLR проводили в трех экземплярах. Индекс пролиферации рассчитывали с использованием программной платформы FlowJo. Трансфекция ДНКазой ICD14 + РВМС трансфицировали 1,5 мг ДНКазы I (Roche) или инактивированной нагреванием ДНКазы I (75 ° C в течение 10 мин / ед.) С использованием реагента для трансфекции DOTAP (Roche) в соответствии с инструкциями производителя. После инкубации с DOTAP-ДНКазой I или DOTAP-HI-DNase I в течение 3 часов (смесь для трансфекции добавляли в культуральную среду), клетки промывали для удаления избытка реагента трансфекции и фермента, стимулировали IFNα и инкубировали в течение 18 часов или GMCSF 100 нг / мл и IFNα 400 нг / мл или IL4 20 нг / мл и инкубировали в течение 3 дней.Маркеры HLA-DR и CD86 анализировали методом проточной цитометрии. Электропорация первичных моноцитов с миРНКCD14 + PBMC были выделены из образцов крови не старше 2 часов и инкубированы в течение 1 часа в RPMI-1640, содержащем 10% FBS, стрептомицин 100 мкг / мл, пенициллин 100 Ед / мл, глутамин. 2 мМ и 1 мМ пирувата натрия в увлажненном инкубаторе 37 ° C / 5% CO 2 перед электропорацией. 10 6 клеток / образец подвергали электропорации с помощью 90 нМ (3 × 30 нМ) дуплексов миРНК Stealth RNAi с использованием набора Amaxa Human Dendritic Cell Nucleofector Kit.Клетки инкубировали со смесью для трансфекции в среде в течение 6 часов, а затем промывали и инкубировали в течение 3 дней с GMCSF и IFNα или IL4. Все эксперименты по трансфекции проводили в трех повторностях (биологические повторы) для всех условий. Эффективность трансфекции оценивали с помощью проточной цитометрии для GFP в клетках, трансфицированных 5 мкг pmaxGFP® Vector (эффективность 65–75%). Подготовка библиотеки для секвенирования РНКРНК была выделена из CD14 + PBMC 7 пациентов и 5 здоровых людей из контрольной группы.Библиотеки секвенирования РНК получали с помощью набора TruSeq RNA, используя 1 мкг общей РНК в соответствии с протоколами Illumina. Все 12 образцов были объединены и смешаны в равных количествах. Образцы были секвенированы в Греческом геномном центре (GGC) при BRFAA. Чтения с парным концом 40bp были произведены с помощью NextSeq500. Анализ библиотеки секвенирования РНКНеобработанные данные секвенирования были сопоставлены с геномом человека GRCh47 / hg19 с использованием tophat2 (версия 2.0.9) (Kim et al., 2013) и параметра «–b2-very-sensitive».Samtools (версия 0.1.19) (Li et al., 2009) использовались для фильтрации данных и преобразования формата файлов, в то время как счетчик HT-seq (версия 0.6.1p1) (Anders et al., 2015) использовался для назначения выровненных фрагментов в экзоны с помощью команды «-s –no –m пересечение –nonempty». Дифференциальную экспрессию выполняли с помощью пакета DESeq R (Anders and Huber, 2010), и гены со значением кратности изменения 1,5 и значением p ≤ 0,05 считались дифференциально экспрессируемыми (DEG) между 5 здоровыми и 7 пациентами (Таблица S3).Онтология генов и анализ путей проводились в DEG с помощью базы знаний DAVID (Huang et al., 2009). Данные были депонированы в архиве краткого чтения (SRA) в рамках BioProject PRJNA392602. Приготовление кислотных наночастиц (aNP)Резомер ® RG 503H PLGA (Sigma-Aldrich, 719870) использовали с соотношением лактид-гликозид 50:50, молекулярная масса от 24000 до 38000. Готовили исходный раствор полимера. растворением 31 мг PLGA в 3,1 мл THF (Sigma-Aldrich, 401757).200 мкл этого исходного раствора добавляли к 20 мл деионизированной воды при обработке ультразвуком. Затем суспензию PLGA-aNP медленно концентрировали, чтобы получить образец с типичной концентрацией 0,18 мг / мл. Для всех экспериментов использовали свежеприготовленные растворы PLGA-aNP. ИммуноблоттингКлетки лизировали в буфере RIPA (50 мМ Трис · HCl при pH 8, 150 мМ NaCl, 0,5% дезоксихолат натрия, 1% Nonidet P-40 и 0,1% SDS) с добавлением ингибиторов протеаз и фосфатаз (Complete EDTA Free; Roche Applied Science) и равные количества белков подвергали SDS-PAGE в 12% гелях и затем переносили на мембрану Immobilon-Psq (Millipore).Мембраны блокировали 5% обезжиренным молоком, 1% BSA в TBST, а затем инкубировали с анти-LC3B, анти-TOMM20 и анти-Beclin1 от Novus Biologicals, анти-SQSTM-1 с помощью MBL, анти-VDAC1 и анти-актин от Millipore. , анти-Stat1 и анти-pStat1 от BD Biosciences, анти-pmTOR, анти-mTOR, анти-ULK1 и анти-pULK1 от Cell Signaling, анти-pBeclin1 от Abbiotec, антимышиный HRP от Milipore и анти-кроличий HRP от Cell Сигнализация согласно предложениям производителей. В качестве контрольного образца использовали белковый экстракт из линии клеток Neuro 2A.Изображение было разрешено системой ECL (Pierce и Millipore) и обнаружено ImageBlot (BIORAD). Относительная интенсивность полос рассчитывалась с помощью программного обеспечения Fiji. Иммунофлуоресценция (IF)Для микроскопии IF клетки высевали на покровные стекла поли-L-лизина (1,5-2 × 10 5 клеток / слайд). Когда соответствующие обработки были закончены, предметные стекла промывали один раз PBS, фиксировали 4% PFA в течение 20 минут (RT), повторно промывали дважды PBS, повторно фиксировали и пропитывали 100% ледяным MetOH в течение 10 минут (RT) и блокировали 2%. BSA в 0,1% сапонине (BS).Первичные антитела оставляли на 1 час при комнатной температуре или O / N при 4 ° C. Срезы несколько раз промывали BS, а вторичные антитела инкубировали в течение 1 часа при комнатной температуре. Используемые антитела: мышиные анти-LC3B (1:20, 5F10 nanoTools), крысиные анти-Lamp-1 (1: 400, 1D4B Abcam), кроличьи антитела против p62 (1: 500, MBL), кроличьи антитела против TFEB (1: 100, 954604 R&D Systems) с последующей инкубацией с Alexa fluor® 555 против мышиных IgG (1: 500, молекулярные зонды), Alexa fluor® 488 против кроличьих IgG (1: 500, молекулярные зонды), Alexa fluor® 633 против IgG крысы (1: 250, молекулярные зонды).Для визуализации ядер использовался DAPI (Sigma-Aldrich). Образцы были закреплены с помощью Mowiol и визуализированы с использованием инвертированной конфокальной системы визуализации живых клеток Leica SP8. Для окрашивания Mitotracker CMXRos и Lysotracker DND-99 (Molecular Probes) красители добавляли в культуру, как рекомендовано производителем, промывали, фиксировали 4% PFA в течение 20 минут (RT) и устанавливали для немедленного наблюдения под микроскопом. Анализ конфокальной визуализацииИспользуемый метод количественной оценки% колокализации основан на финансируемом NIH макросе, разработанном Дэниелом Дж.Shiwarski, B.S., University of Pittsburgh, и подтверждено для анализа аутофагического потока Pampliega et al. (2013). Этот сценарий (макрос) был запущен в программном обеспечении Фиджи, и числовые результаты выражали степень совместной локализации зеленого и красного точек в изображениях RGB. TaqMan AssayДНКвыделяли с помощью набора QIAamp DNA Micro (QIAGEN) в соответствии с протоколом производителя. Для количественного определения количества копий митохондриальной ДНК использовали мультиплексный анализ ОТ-ПЦР в соответствии с Phillips et al.(2014). Образцы ДНК (50 нг / реакция) использовали вместе с соответствующими объемами TaqMan Fast Universal PCR Master Mix (2x), No AmpErase UNG (Applied Biosystems)) в системе реального времени CFX ConnectTM. Общий объем каждой мультиплексной реакции RT-PCR составлял 25 мкл. huβ2Μ праймеры — F: TTAACGTCCTTGGCTGGGTC, Р: ACTGGAAGACAAAGGGCTCG, huβ2Μ зонд: CAGATGCAGTCCAAACT-CTCACT, mtMinArc праймеры — F: CTGTTCCCCAACCTTTTCCT, Р: CCATGATTGT-GAGGGGTAGG, mtMinArc зонд: GACCCCCTAACAACCCCC СТИНГ (TMEM173) праймеры и зонды: Hs00736958_m1 анализ, GAPDH праймеры и зонды : Анализ Hs02758991_g1 и праймеры и зонд TLR9: анализ Hs00152973_m1 от Applied Biosystems. ПЦР в реальном времени (RT-PCR)Общую РНК из моноцитов собирали с использованием протокола экстракции TRIzol (Invitrogen) с обработкой Turbo DNase (Ambion) в соответствии с инструкциями производителя. (- ΔΔCT)]. Последовательности праймеров использовали: IFN & alpha; F: GGTGACAGAGACTCCCCTGA, R: CAGGCACAAGGGCTGTATTTCTT, CXCL10 — F: GTGGCATTCAAGGAGTACCTC, R: TGATGGCCTTCGATTCTGGATT, LC3B-F: CTGTTGGTGAACGGACACAG, R: CTGGGAGGCATAGACCATGT, ATG5-F: TGACGTTGGTAACTGACAAAGTG, R: AATGCCATTTCAGTGGTGTG, P62 / SQSTM1- F: AGCAGATGAGGAAGATCGCC, Р: CTGTAGACGGGTCCACTTCTT, PINK1-F: GGATATGGAGCAGTCA CTTACAG, Р: GGCAGCACATCAGGGTAGTC, PARKIN- F: CGACCCTCAACTT GGCTACT Р: TCTTTAATCAAGGAGTTGGGACA, TLR9 — F: CTGCCTTCCTACCCTGTGAG, Р: GGATGCGGTTGGAGGACAA, GAPDH — F: CATGTTCCAATATGATTCCACC, Р: GATGGG ATTTCCATTGATGAC. ELISAОбнаружение человеческого IL-6, CXCL10, TNFα (eBioscience) и IFNα (VeriKine Human Interferon Alpha Multi-Subtype Serum ELISA Kit, PBL) в сыворотке человека и супернатантах культур, собранных в указанные моменты времени, выполняли с помощью сэндвич-ИФА в соответствии с рекомендациями производителя. Светопоглощение измеряли с помощью планшет-ридера ELx800 Biotek. Фоновый сигнал был вычтен. Все образцы оценивались в двух экземплярах (технические повторности). Анализ АТФМоноциты CD14 + (10 6 клеток / состояние) собирали, промывали и лизировали в соответствии с протоколом производителя (Abcam) для колориметрического анализа.Светопоглощение измеряли с помощью планшет-ридера ELx800 Biotek. Уровни АТФ рассчитывали по следующей формуле: [ ATP ] ( нмоль на мкл или мМ ) = ( Ts / Sv ) D, Ts = количество АТФ из стандартной кривой (нмоль или мМ) , Sv = объем образца, добавленный в лунки для образцов (мкл), D = коэффициент разбавления образца. Выделение митохондрийМитохондрии выделяли из культивируемых клеток с помощью набора для выделения митохондрий, Thermo Fisher Scientific (89874), в соответствии с протоколом производителя.Вкратце, клетки лизировали с использованием метода на основе реагентов в соответствии с инструкциями для получения более очищенной фракции митохондрий с> 50% снижением лизосомных и пероксисомальных примесей. Для вестерн-блот-анализа осадок митохондрий кипятили с буфером для образцов SDS-PAGE: буфером RIPA с добавлением ингибиторов фосфатазы и протеаз (ThermoFisher Scientific) и дополнительным SDS (20%) и β-меркаптоэтанолом (5%). Определение активности катепсина DАктивность катепсина D измеряли с помощью набора для флуорометрического анализа от Abcam (ab65302) в соответствии с инструкциями производителя.Вкратце, 10 6 клеток собирали для каждого состояния, немедленно лизировали и инкубировали с предпочтительным субстратом катепсина-D GKPILFFRLK (Dnp) -D-R-Nh3, меченным MCA. Флуоресценцию, высвобождаемую при расщеплении субстрата, измеряли на флюоресцентном микропланшет-ридере FLx800. Для всех экспериментов клеточные лизаты оценивали как свежеприготовленные. Измерения скорости потребления кислородаИзмерения скорости потребления кислорода проводились, как описано Palikaras et.al. на выставке Nature 2015 с использованием электрода Кларка (Hansatech Instruments).4×10 6 CD14 + моноцитов, обработанных, как указано, промывали и собирали в буфере PBS. В камере поддерживалась температура 37 ° C, измерения проводились в течение 10–15 мин в зависимости от скорости потребления кислорода. Наклон прямых участков графиков использовался для расчета скорости потребления кислорода. Клетки извлекали после измерения дыхания и собирали для количественного определения белка. Нормы были нормализованы по содержанию белка, рассчитанному с помощью анализа белка Брэдфорда. РеагентыПлаквенил (CQ: 46.1 мкМ) был приобретен у Санофи Авентис. Рапамицин (Rapam .: 1 мкМ) был от SIGMA. Вортманин (WM: 1 мкМ) был приобретен у InvivoGen. Mitotracker CMXRos (MTR: 1 мкМ), MitoTEMPO (2 нМ), Lysotracker-DND99 (LTR: 1 мкМ) и MitoSOX (5 мкл) были приобретены у компании Molecular Probes — Thermo Fisher Scientific. JC-1 (2 мкг / мл) был приобретен в eBiosciences. Рекомбинантная ДНКаза I и реагент липосомальной трансфекции DOTAP были получены от Roche. Человеческий STING (TMEM173), TLR9 и дуплексные олигонуклеотиды Negative Stealth siRNA были приобретены у Invitrogen. Количественная оценка и статистический анализСтатистический анализРезультаты представлены как среднее значение ± стандартная ошибка среднего. Статистический анализ проводился с использованием парного критерия Стьюдента или непарного непараметрического U-критерия Манна-Уитни, как указано в легенде каждого рисунка. Все эксперименты проводили в количестве биологических повторов, указанном как n в каждой легенде к рисункам. Вкратце, эксперименты, проведенные на разных здоровых донорах (n соответствует количеству здоровых доноров), были разработаны для оценки эффекта различных методов лечения, оценивающих одного донора / эксперимент.В этом случае статистический анализ проводился с использованием парного t-критерия, в то время как сравнение здоровых образцов с образцами SLE было статистически проанализировано с использованием непарного t-критерия Стьюдента (непараметрический критерий Манна-Уитни). Значение P <0,05 считалось показателем статистической значимости ( ). Все данные были проанализированы с помощью программного обеспечения GraphPad Prism. БлагодарностиМы благодарим всех пациентов, предоставивших клинические образцы, С. Георгакиса за помощь в проведении экспериментов, А. Баноса за помощь в подготовке образцов для RNA-seq, К.Паликарасу за помощь в анализе потребления кислорода и И. Гергианнаки за организацию историй болезни пациентов с СКВ, лечившихся в ревматологической клинике университетской больницы Крита. Эта работа была выполнена Греческим Генеральным секретариатом исследований и технологий (софинансируется Европейским социальным фондом [ESF] и Национальными ресурсами) Aristeia I 2344, а также Фондом исследований в области ревматологии (FOREUM). КГ. был профинансирован IKY-Siemens (Стипендии передового опыта после докторов наук), а Д.Б. и П.В. были профинансированы Проектом инновационной медицины Европейского союза 6 (контракт «BeTheCure» 115142-2) и Европейским исследовательским советом (ERC) в рамках исследовательской и инновационной программы Европейского союза Horizon 2020 (соглашение о гранте 742390). Эта работа также была поддержана COST Actions Mye-EUNITER (BM1404, http://www.mye-euniter.eu) и TRANSAUTOPHAGY (CA15138, http://cost-transautophagy.eu), которые являются частью Рамочной программы Европейского Союза. Horizon 2020. Вклад авторовКонцептуализация и методология, К.ГРАММ.; Расследование, K.G., E.K., D.N., A.P., and A.B .; Письмо — Оригинал, К.Г .; Writing — Review & Editing, G.B., D.T.B., and P.V .; Ресурсы, A.M., P.S., G.B., D.T.B., and P.V .; Финансирование приобретения, K.G., G.B., P.S., D.T.B., and P.V .; Надзор, D.T.B. и П.В. Декларация интересовАвторы заявляют об отсутствии конкурирующих интересов. ПримечанияДата публикации: 23 октября 2018 г. Дополнительная информацияДокумент S1. Рисунки S1 – S6 и таблицы S2 и S4: Таблица S1.Клинические записи пациентов с СКВ, относящиеся к рисунку 1: Таблица S3. RNA-Seq DEG, относящиеся к фиг. 6A:Таблица, содержащая 380 дифференциально экспрессируемых генов (DEG) между пациентом и группой здоровых людей. Документ S2. Статья плюс дополнительная информация:СсылкиAlissafi T., Banos A., Boon L., Sparwasser T., Ghigo A., Wing K., Vassilopoulos D., Boumpas D., Chavakis T., Cadwell K., Verginis P. Tregs сдерживают аутофагию дендритных клеток, чтобы улучшить аутоиммунитет.J. Clin. Вкладывать деньги. 2017; 127: 2789–2804. [Бесплатная статья PMC] [PubMed] [Ученый Google] Андерс С., Пил П.Т., Хубер В. HTSeq — среда Python для работы с данными высокопроизводительного секвенирования. Биоинформатика. 2015; 31: 166–169. [Бесплатная статья PMC] [PubMed] [Google Scholar] Бланко П., Палука А.К., Гилл М., Паскуаль В., Баншеро Дж. Индукция дифференцировки дендритных клеток с помощью IFN-альфа при системной красной волчанке. Наука. 2001; 294: 1540–1543. [PubMed] [Google Scholar] Бурденкс М., Дэниел Дж., Генин Э., Сориа Ф.N., Blanchard-Desce M., Bezard E., Dehay B. Наночастицы восстанавливают лизосомные дефекты закисления: последствия для болезни Паркинсона и других заболеваний, связанных с лизосомами. Аутофагия. 2016; 12: 472–483. [Бесплатная статья PMC] [PubMed] [Google Scholar] Bride KL, Vincent T., Smith-Whitley K., Lambert MP, Bleesing JJ, Seif AE, Manno CS, Casper J., Grupp SA, Teachey D.T. Сиролимус эффективен при рецидивирующие / рефрактерные аутоиммунные цитопении: результаты проспективного мультиинституционального исследования. Кровь. 2016; 127: 17–28.[Бесплатная статья PMC] [PubMed] [Ученый Google] Кайелли С., Атал С., Домик Б., Мюрат Э., Чандра М., Банчеро Р., Байш Дж., Фелпс К., Клейтон С., Гонг М. Окисленные митохондриальные нуклеоиды, высвобождаемые нейтрофилами, приводят к выработке интерферона I типа при волчанке человека. J. Exp. Med. 2016; 213: 697–713. [Бесплатная статья PMC] [PubMed] [Google Scholar] Канауд Г., Бьенайме Ф., Табарин Ф., Батайон Г., Сейлхин Д., Ноэль Л.-Х., Драгон-Дюрей М.-А., Снанудж Р. ., Фридлендер Г., Хальбвакс-Мекарелли Л. Ингибирование пути mTORC при антифосфолипидном синдроме.N. Engl. J. Med. 2014; 371: 303–312. [PubMed] [Google Scholar] Цан К., Чжоу Ю., Наварро Б., Со Й.-Дж., Аранда К., Ши Л., Батталья-Сю С., Ниссим И., Клэпхэм Д.Э., Рен Д. mTOR регулирует лизосомные АТФ-чувствительные двухпоровые Na (+) каналы для адаптации к метаболическому состоянию. Клетка. 2013; 152: 778–790. [Бесплатная статья PMC] [PubMed] [Google Scholar] Chen J., Fang Y. Новый путь, регулирующий передачу сигналов рапамицина (mTOR) млекопитающим. Biochem. Pharmacol. 2002; 64: 1071–1077. [PubMed] [Google Scholar] Чен К., Liu Y., Liu R., Ikenoue T., Guan K.-L., Liu Y., Zheng P. TSC-mTOR поддерживает покой и функцию гемопоэтических стволовых клеток путем подавления митохондриального биогенеза и активных форм кислорода. J. Exp. Med. 2008; 205: 2397–2408. [Бесплатная статья PMC] [PubMed] [Google Scholar] Кроу Ю.Дж., Синдром Манеля Н. Айкарди-Гутьера и интерферонопатии типа I. Nat. Rev. Immunol. 2015; 15: 429–440. [PubMed] [Google Scholar] de Jong SD, Basha G., Wilson KD, Kazem M., Cullis P., Jefferies W., Tam Y. Иммуностимулирующая активность неметилированного и метилированного олигодезоксинуклеотида CpG зависит от их способности колокализоваться с TLR9 в поздних эндосомах.J. Immunol. 2010. 184: 6092–6102. [PubMed] [Google Scholar] Динг Д., Мехта Х., МакКьюн У. Дж., Каплан М. Дж. Аберрантный фенотип и функция миелоидных дендритных клеток при системной красной волчанке. J. Immunol. 2006; 177: 5878–5889. [PubMed] [Google Scholar] Гейслер С., Хольмстрём К.М., Скуят Д., Физель Ф.С., Ротфус О.С., Кале П.Дж., Спрингер В. PINK1 / Паркин-опосредованная митофагия зависит от VDAC1 и p62 / SQSTM1. Nat. Cell Biol. 2010. 12: 119–131. [PubMed] [Google Scholar] Гергей П., младший, Ниланд Б., Гончоров Н., Pullmann R., Jr., Phillips P.E., Perl A. Устойчивая гиперполяризация митохондрий, повышенная продукция реактивного промежуточного кислорода и подщелачивание цитоплазмы характеризуют измененную передачу сигналов IL-10 у пациентов с системной красной волчанкой. J. Immunol. 2002; 169: 1092–1101. [Бесплатная статья PMC] [PubMed] [Google Scholar] Гергели П., младший, Гроссман К., Ниланд Б., Пушкаш Ф., Неупане Х., Аллам Ф., Банки К., Филлипс П. Е., Перл А. Митохондриальные гиперполяризация и истощение АТФ у пациентов с системной красной волчанкой.Rheum артрита. 2002; 46: 175–190. [Бесплатная статья PMC] [PubMed] [Google Scholar] Gujer C., Sandgren KJ, Douagi I., Adams WC, Sundling C., Smed-Sörensen A., Seder RA, Karlsson Hedestam GB, Loré K. Произведен IFN-α человеческими плазматическими дендритными клетками усиливает Т-клеточную дифференцировку наивных В-клеток. J. Leukoc. Биол. 2011; 89: 811–821. [Бесплатная статья PMC] [PubMed] [Google Scholar] Хуанг В., Шерман Б.Т., Лемпицки Р.А. Систематический и комплексный анализ больших списков генов с использованием ресурсов биоинформатики DAVID.Nat. Protoc. 2009. 4: 44–57. [PubMed] [Google Scholar] Егерленер А., Маурер П., Бесса Дж., Хинтон Х. Дж., Копф М., Бахманн М. Ф. Передача сигнала TLR9 в В-клетках определяет рекомбинацию переключения класса на IgG2a. J. Immunol. 2007; 178: 2415–2420. [PubMed] [Google Scholar] Ким Д., Пертя Г., Трапнелл К., Пиментел Х., Келли Р., Зальцберг С.Л. TopHat2: точное выравнивание транскриптомов при наличии вставок, делеций и слияний генов. Genome Biol. 2013; 14: R36. [Бесплатная статья PMC] [PubMed] [Google Scholar] Клинман Д.М., Йи А.К., Бокаж С.Л., Коновер Дж., Криг А.М. Мотивы CpG, присутствующие в ДНК бактерий, быстро побуждают лимфоциты секретировать интерлейкин 6, интерлейкин 12 и интерферон гамма. Proc. Natl. Акад. Sci. США. 1996; 93: 2879–2883. [Бесплатная статья PMC] [PubMed] [Google Scholar] Клионски Д. Дж., Бэреке Э. Х., Брумелл Дж. Х., Чу С. Т., Кодоньо П., Куэрво А. М., Дебнат Дж., Деретич В., Элазар З., Эскелинен Э.-Л. Исчерпывающий глоссарий молекул и процессов, связанных с аутофагией (2-е издание) Аутофагия. 2011; 7: 1273–1294.[Бесплатная статья PMC] [PubMed] [Google Scholar] Клионски Д.Д., Абдельмохсен К., Абэ А., Абедин М.Дж., Абелиович Х., Асеведо Арозена А., Адачи Х., Адамс К.М., Адамс П.Д., Адели К. Рекомендации для использование и интерпретация анализов для мониторинга аутофагии (3-е издание) Аутофагия. 2016; 12: 1–222. [Бесплатная статья PMC] [PubMed] [Google Scholar] Корольчук В.И., Сайки С., Лихтенберг М., Сиддики Ф.Х., Робертс Э.А., Имарисио С., Джахрейсс Л., Саркар С., Футтер М., Мензис Ф.М. Позиционирование лизосом координирует реакцию клеток на питательные вещества.Nat. Cell Biol. 2011; 13: 453–460. [Бесплатная статья PMC] [PubMed] [Google Scholar] Крыско Д.В., Агостинис П., Крыско О., Гарг А.Д., Бахерт К., Ламбрехт Б.Н., Ванденабеле П. Возникающая роль связанных с повреждениями молекулярных паттернов, полученных из митохондрий, в воспалении. Trends Immunol. 2011. 32: 157–164. [PubMed] [Google Scholar] Ли Х., Хандакер Б., Вайсокер А., Феннелл Т., Руан Дж., Гомер Н., Март Г., Абекасис Г., Дурбин Р., Подгруппа обработки данных проекта «1000 геномов» The Выравнивание последовательности / формат карты и SAMtools.Биоинформатика. 2009. 25: 2078–2079. [Бесплатная статья PMC] [PubMed] [Google Scholar] Liao X., Reihl A.M., Luo X.M. Нарушение иммунной толерантности при системной красной волчанке дендритными клетками. J. Immunol. Res. 2016; 2016: 6269157. [Бесплатная статья PMC] [PubMed] [Google Scholar] Ма Ю., Галлуцци Л., Зитвогель Л., Кремер Г. Аутофагия и клеточные иммунные реакции. Иммунитет. 2013; 39: 211–227. [PubMed] [Google Scholar] Ма З., Джейкобс С. Р., Вест Дж. А., Стопфорд К., Чжан З., Дэвис З., Барбер Г. Н., Глаунсингер Б.А., Диттмер Д.П., Дамания Б. Модуляция пути зондирования ДНК cGAS-STING гамма-герпесвирусами. Proc. Natl. Акад. Sci. США. 2015; 112: E4306 – E4315. [Бесплатная статья PMC] [PubMed] [Ученый Google] Мацумото А., Итикава Т., Накао К., Мияаки Х., Хирано К., Фудзимито М., Акияма М., Миума С., Одзава Э., Сибата Х. Индуцированная интерфероном-α активация mTOR является сигналом вируса гепатита С через фосфатидилинозитол-3-киназу-Akt-независимый путь. J. Gastroenterol. 2009. 44: 856–863. [PubMed] [Google Scholar] Минтон К.Инфламмасома: противовоспалительный эффект митофагии. Nat. Rev. Immunol. 2016; 16 206–206. [PubMed] [Google Scholar] Накай А., Ямагути О., Такеда Т., Хигучи Ю., Хикосо С., Таниике М., Омия С., Мизоте И., Мацумура Ю., Асахи М. Роль аутофагии в кардиомиоцитах в базальном состоянии и в ответ на гемодинамический стресс. Nat. Med. 2007. 13: 619–624. [PubMed] [Google Scholar] Ниволд Т. Б., Хуа Дж., Леман Т. Дж. А., Харли Дж. Б., Кроу М. К. Высокая активность IFN-α в сыворотке крови является наследственным фактором риска системной красной волчанки.Genes Immun. 2007. 8: 492–502. [Бесплатная статья PMC] [PubMed] [Google Scholar] Оукс З., Винанс Т., Каза Т., Фернандес Д., Лю Ю., Ландас С.К., Банки К., Перл А. Дисфункция митохондрий в печени и антифосфолипидные антитела продуцирование предшествует началу заболевания и реагирует на рапамицин у склонных к волчанке мышей. Arthritis Rheumatol. 2016; 68: 2728–2739. [Бесплатная статья PMC] [PubMed] [Ученый Google] Ока Т., Хикосо С., Ямагути О., Танейке М., Такеда Т., Тамай Т., Оябу Дж., Муракава Т., Накаяма Х., Нисида К. .Митохондриальная ДНК, ускользающая от аутофагии, вызывает воспаление и сердечную недостаточность. Природа. 2012; 485: 251–255. [Бесплатная статья PMC] [PubMed] [Google Scholar] Pampliega O., Orhon I., Patel B., Sridhar S., Díaz-Carretero A., Beau I., Codogno P., Satir BH, Satir P., Cuervo ЯВЛЯЮСЬ Функциональное взаимодействие аутофагии и цилиогенеза. Природа. 2013; 502: 194–200. [Бесплатная статья PMC] [PubMed] [Google Scholar] Паскуаль В., Фаркас Л., Баншеро Дж. Системная красная волчанка: все пути ведут к интерферонам I типа.Curr. Opin. Иммунол. 2006. 18: 676–682. [PubMed] [Google Scholar] Пирс Э.Л., Поффенбергер М.С., Чанг С.-Х., Джонс Р.Г. Подпитка иммунитета: понимание метаболизма и функции лимфоцитов. Наука. 2013; 342: 1242454. [Бесплатная статья PMC] [PubMed] [Google Scholar] Филлипс Н.Р., Спроус М.Л., Роби Р.К. Одновременная количественная оценка количества копий митохондриальной ДНК и степени делеций: мультиплексный анализ ПЦР в реальном времени. Sci. Отчет 2014; 4: 3887. [Бесплатная статья PMC] [PubMed] [Google Scholar] Писецкий Д.С. Роль митохондрий в иммуноопосредованных заболеваниях: опасности раздвоения личности.Arthritis Res. Ther. 2016; 18: 169. [Бесплатная статья PMC] [PubMed] [Google Scholar] Рави С., Митчелл Т., Крамер П., Чако Б., Дарли-Усмар В.М. Митохондрии в моноцитах и макрофагах — значение для трансляционных и фундаментальных исследований. Int. J. Biochem. Cell Biol. 2014; 53: 202–207. [Бесплатная статья PMC] [PubMed] [Ученый Google] Рейман Дж., Брагонци А., Конезе М. Роль эндоцитоза, опосредованного клатрином и кавеолами, в переносе генов, опосредованном липо- и полиплексами. Мол. Ther. 2005; 12: 468–474. [PubMed] [Google Scholar] Шмайссер Х., Фей С.Б., Хоровиц Дж., Фишер Э. Р., Балински К. А., Мияке К., Бекиш Дж., Сноу А. Л., Зун К. Интерфероны типа I вызывают аутофагию в некоторых линиях раковых клеток человека. Аутофагия. 2013; 9: 683–696. [Бесплатная статья PMC] [PubMed] [Google Scholar] Сена Л.А., Ли С., Джайраман А., Пракрия М., Эзпонда Т., Хильдеман Д.А., Ван С.-Р., Шумакер П.Т., Лихт Д.Д., Перлман Х. Митохондрии необходимы для активации антиген-специфических Т-клеток посредством передачи сигналов активных форм кислорода. Иммунитет. 2013. 38: 225–236. [Бесплатная статья PMC] [PubMed] [Google Scholar] Сеттембр К., Ди Мальта К., Полито В.А., Аренсибия М.Г., Ветрини Ф., Эрдин С., Эрдин С.Ю., Хьюнх Т., Медина Д., Колелла П. TFEB связывает аутофагию с биогенезом лизосом. Наука. 2011; 332: 1429–1433. [Бесплатная статья PMC] [PubMed] [Google Scholar] Шривастав М., Невольд Т. Датчики нуклеиновых кислот и продукция интерферона типа I при системной красной волчанке. Фронт. Иммунол. 2013; 4: 319. [Бесплатная статья PMC] [PubMed] [Google Scholar] Сухбаатар Н., Хенгстшлегер М., Вейхарт Т. Регуляция дифференцировки и функции дендритных клеток, опосредованная mTOR.Trends Immunol. 2016; 37: 778–789. [Бесплатная статья PMC] [PubMed] [Google Scholar] Цокос Г.К. Системная красная волчанка в 2015 г .: клеточные и метаболические потребности эффекторных Т-клеток. Nat. Rev. Rheumatol. 2016; 12: 74–76. [PubMed] [Google Scholar] Уол Д. Р., Байерсдорфер С. А., Феррара Дж. Л. М., Опипари А. В., мл., Глик Г. Д. Отдельные метаболические программы в активированных Т-клетках: возможности для селективной иммуномодуляции. Иммунол. Ред. 2012; 249: 104–115. [Бесплатная статья PMC] [PubMed] [Google Scholar] Weckerle C.Э., Франек Б.С., Келли Дж. А., Кумабе М., Миколайтис Р.А., Грин С.Л., Утсет Т.О., Джолли М., Джеймс Дж. А., Харли Дж. Б., Неволд Т. Б. Сетевой анализ ассоциаций между активностью интерферона-α в сыворотке крови, аутоантителами и клиническими особенностями системной красной волчанки. Rheum артрита. 2011; 63: 1044–1053. [Бесплатная статья PMC] [PubMed] [Google Scholar] West A.P., Khoury-Hanold W., Staron M., Tal M.C., Pineda C.M., Lang S.M., Bestwick M., Duguay B.A., Raimundo N., MacDuff D.A. Стресс митохондриальной ДНК стимулирует противовирусный врожденный иммунный ответ.Природа. 2015; 520: 553–557. [Бесплатная статья PMC] [PubMed] [Google Scholar] Yin Y., Choi S.-C., Xu Z., Perry DJ, Seay H., Croker BP, Sobel ES, Brusko TM, Morel L. Нормализация CD4 + T клеточный метаболизм обращает волчанку вспять. Sci. Пер. Med. 2015; 7: 274ра18. [Бесплатная статья PMC] [PubMed] [Google Scholar] Zhang Q., Raoof M., Chen Y., Sumi Y., Sursal T., Junger W., Brohi K., Itagaki K., Hauser CJ. Циркулирующие митохондриальные DAMPs вызывают воспалительные реакции на травму. Природа. 2010; 464: 104–107. [Бесплатная статья PMC] [PubMed] [Google Scholar] Zhang Y., Морган М.Дж., Чен К., Чокси С., Лю З.Г. Индукция аутофагии необходима для дифференцировки моноцитов и макрофагов. Кровь. 2012; 119: 2895–2905. [Бесплатная статья PMC] [PubMed] [Google Scholar] Zhang YM, Cheng FJ, Zhou XJ, Qi YY, Hou P., Zhao MH, Zhang H. Выявление генетических ассоциаций между ATG5 и волчаночным нефритом с помощью trans -eQTL . J. Immunol. Res. 2015; 2015: 153132. [Бесплатная статья PMC] [PubMed] [Google Scholar] Чжоу X.J., Lu X.L., Lv J.C., Yang H.Z., Qin L.X., Чжао М.Х., Су Ю., Ли З.Г., Чжан Х. Генетическая ассоциация межгенной области PRDM1-ATG5 и аутофагия с системной красной волчанкой в китайской популяции. Аня. Реум. Дис. 2011; 70: 1330–1337. [PubMed] [Google Scholar] Zhu S., Qiao Y., Wu J., Zang G., Liu Y.-J., Chen J. Раковые клетки человека реагируют на цитозольные нуклеиновые кислоты через усиленное производство интерферона-β и апоптоз . J. Immunother. Рак. 2015; 3: 250. [Google Scholar]Произошла ошибка при настройке пользовательского файла cookieЭтот сайт использует файлы cookie для повышения производительности.Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт. Настройка вашего браузера для приема файлов cookieСуществует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
Почему этому сайту требуются файлы cookie?Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня. Что сохраняется в файле cookie?Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется. Как правило, в файлах cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать. .About the author |