Серия, тип | Краткая техническая характеристика | Обозначение ТУ, ТЗ | |
ППК-8042 | Номинальное напряжение, В | 900 | ТУ 16-526.353-74 |
Род тока | постоянный | ||
Номинальный ток, А | |||
ППК-8023, 8023М, 8042, 8063, 8063М, 8064, 8064М | 1000 | ||
900 | |||
Номинальное напряжение катушки вентиля пневматического привода, В | |||
ППК-8023 | 24 | ||
ППК-8023, 8023М, 8063, 8063М | 75 | ||
ППК-8042, 8064, 8064М, 8122, 8122М | 110 | ||
Номинальное давление, МПа | 0,5 | ||
Максимальное давление, МПа | 0,675 | ||
Минимальное давление, МПа | 0,35 | ||
Масса, кг | |||
ППК-8023, 8023М | 106 | ||
ППК-8042 | 132 | ||
ППК-8063, 8063М, 8064, 8064М, 8122, 8122М | 152 | ||
Климатическое исполнение | УЗ, ТЗ | ||
ППК-12201 | Род тока | постоянный | ТУ 16-526. |
Номинальный ток, А | |||
Номинальный напряжение, В | 1000 | ||
Номинальное напряжение катушки вентиля пневматического привода, В: | |||
ППК-12201 | 75 | ||
ППК-12602, ППК-12062, ППК-12802, ППК-12804, ППК-12084 | 110 | ||
Номинальное давление, МПа | 0,5 | ||
Максимальное давление, МПа | 0,675 | ||
Минимальное давление, МПа | 0,35 | ||
Масса, кг | |||
| ППК-12201 | 50 | ||
ППК-12602, 12062 | 95 | ||
ППК-12802, 12082, 12804, 12084 | 105 | ||
Климатическое исполнение | УЗ, УХЛЗ, ТЗ | ||
ППК-11201 | Номинальный ток, А | 800 | ТУ 16-526. |
Номинальное напряжение катушки вентиля пневматического привода, В: | |||
ППК-11201 | |||
ППК-11602 | 110 | ||
Масса, кг | |||
ППК-11201 | 45 | ||
ППК-11602 | 87 | ||
 510-83
510-83 510-83
510-83|
ППК-8023 ППК-8023М ППК-8042 ППК-8042М ППК-8063 ППК-8063М ППК-8064 ППК-8064Д ППК-8122 ППК-8122М |
||
| Контакт главной цепи: | ||
| Номинальное напряжение для серии ППК 8000, В | 900 | |
| Род тока | постоянный | |
| Номинальный ток, А | ||
| ППК-8023, ППК-8023М, ППК-8042, ППК-8063, ППК-8063М, ППК-8064, ППК-8064Д | 1000 | |
| ППК-8122, ППК-8122М | 900 | |
| Контакты вспомогательной цепи: | ||
| Номинальный ток, А | 2 | |
| Номинальное напряжение, В | 110 | |
| Номинальное напряжение катушки вентиля | ||
| пневматического привода, В | ||
| ППК-8023, ППК-8023М | 24,75 | |
| ППК-8063 | 24 | |
| ППК-8063, ППК-8063М | 75 | |
| ППК-8122, ППК-8122М, ППК-8042, ППК-8042М, ППК-8064 | 110 | |
| Номинальное давление воздуха, МПа | 0,5 | |
| Максимальное давление воздуха, МПа | 0,675 | |
| Минимальное давление воздуха, МПа | 0,35 | |
| Масса, кг | ||
| ППК-8023, ППК-8023М | 100 | |
| ППК-8042, ППК-8042М | 132 | |
| ППК-8122, ППК-8122М, ППК-8063, ППК-8063М, ППК-8064, ППК-8064Д | 152 | |
| Климатическое исполнение | У2, Т3 | |
|
ППК12201 ППК12202 ППК12022 ППК12404Д ППК12044Д ППК 12602 ППК12062 ППК12602Д ППК12062Д ППК12802 ППК12082 ППК12804Д ППК12084Д ППК12801 ППК12081 ППК11201 ППК11602 ППК11062 |
Контакты главной цепи: | |
| Номинальное напряжение для серии ППК 12000 | ||
| ППК11000, В | 1000 | |
| Род тока | постоянный | |
| Номинальный ток для серии ППК 12000, А | 1000 | |
| Номинальный ток для серии ППК 11000, А | 800 | |
| Контакты вспомогательной цепи: | ||
| Номинальный ток, А | 2 | |
| Номинальное напряжение катушки вентиля пневматического привода ивспомогательных цепей, В | ||
| Для серии ППК 12000, 11000, В | ||
| ППК 12201, 12801, 12081 | 75 | |
| ППК12202, 12022, 12404Д, 12044Д, 12602, 12062 | 110 | |
| 12602Д, ППК 12062Д, 12082, 12802, 12804Д, 12084Д, 11602, 11062 | ||
| Давление воздуха в приводе для серии переключателей ППК 12000, 11000: | ||
| Номинальное давление воздуха, МПа | 0,5 | |
| Максимальное давление воздуха, МПа | 0,675 | |
| Минимальное давление, МПа | 0,35 | |
| Масса, кг | ||
| ППК12404Д, 12044Д | 70 | |
| ППК 12201, 12202, 12022 | 50 | |
| ППК 12602, 12062, 12602Д, 12062Д | 95 | |
| ППК 12802,12082, 12804Д, 12084Д, 12801, 12081 | 105 | |
| ППК 11201 | 45 | |
| ППК 11602, 11062 | 87 | |
| Климатическое исполнение | У2, УХЛ3, Т3 | |
| ППК 12000, ППК 11000 |
Серия, тип | Краткая техническая информация | |
ППК-8023 ППК-8023М ППК-8042 ППК-8042М ППК-8063 ППК-8063М ППК-8064 ППК-8064Д ППК-8122 ППК-8122М | ||
| Контакт главной цепи: | ||
| Номинальное напряжение для серии ППК 8000, В | 900 | |
| Род тока | постоянный | |
| Номинальный ток, А | ||
| ППК-8023, ППК-8023М, ППК-8042, ППК-8063, ППК-8063М, ППК-8064, ППК-8064Д | 1000 | |
| ППК-8122, ППК-8122М | ||
| Контакты вспомогательной цепи: | ||
| Номинальный ток, А | 2 | |
| Номинальное напряжение, В | 110 | |
| Номинальное напряжение катушки вентиля | ||
| пневматического привода, В | ||
| ППК-8023, ППК-8023М | 24,75 | |
| ППК-8063 | 24 | |
| ППК-8063, ППК-8063М | 75 | |
| ППК-8122, ППК-8122М, ППК-8042, ППК-8042М, ППК-8064 | 110 | |
| Номинальное давление воздуха, МПа | 0,5 | |
| Максимальное давление воздуха, МПа | 0,675 | |
| Минимальное давление воздуха, МПа | 0,35 | |
| Масса, кг | ||
| ППК-8023, ППК-8023М | 100 | |
| ППК-8042, ППК-8042М | 132 | |
| ППК-8122, ППК-8122М, ППК-8063, ППК-8063М, ППК-8064, ППК-8064Д | 152 | |
| Климатическое исполнение | У2, Т3 | |
ППК12201 ППК12202 ППК12022 ППК12404Д ППК12044Д ППК 12602 ППК12062 ППК12602Д ППК12062Д ППК12802 ППК12082 ППК12804Д ППК12084Д ППК12801 ППК12081 ППК11201 ППК11602 ППК11062 | Контакты главной цепи: | |
| Номинальное напряжение для серии ППК 12000 | ||
| ППК11000, В | 1000 | |
| Род тока | постоянный | |
| Номинальный ток для серии ППК 12000, А | 1000 | |
| Номинальный ток для серии ППК 11000, А | 800 | |
| Контакты вспомогательной цепи: | ||
| Номинальный ток, А | 2 | |
| Номинальное напряжение катушки вентиля пневматического привода ивспомогательных цепей, В | ||
| Для серии ППК 12000, 11000, В | ||
| ППК 12201, 12801, 12081 | 75 | |
| ППК12202, 12022, 12404Д, 12044Д, 12602, 12062 | 110 | |
| 12602Д, ППК 12062Д, 12082, 12802, 12804Д, 12084Д, 11602, 11062 | ||
| Давление воздуха в приводе для серии переключателей ППК 12000, 11000: | ||
| Номинальное давление воздуха, МПа | 0,5 | |
| Максимальное давление воздуха, МПа | 0,675 | |
| Минимальное давление, МПа | 0,35 | |
| Масса, кг | ||
| ППК12404Д, 12044Д | 70 | |
| ППК 12201, 12202, 12022 | 50 | |
| ППК 12602, 12062, 12602Д, 12062Д | 95 | |
| ППК 12802,12082, 12804Д, 12084Д, 12801, 12081 | 105 | |
| ППК 11201 | 45 | |
| ППК 11602, 11062 | 87 | |
| Климатическое исполнение | У2, УХЛ3, Т3 | |
| ППК 12000, ППК 11000 | ||
ППК-8023 ППК-8023М ППК-8042 ППК-8042М ППК-8063 ППК-8063М ППК-8064 ППК-8064Д ППК-8122 ППК-8122М | ||
| Контакт главной цепи: | ||
| Номинальное напряжение для серии ППК 8000, В | 900 | |
| Род тока | постоянный | |
| Номинальный ток, А | ||
| ППК-8023, ППК-8023М, ППК-8042, ППК-8063, ППК-8063М, ППК-8064, ППК-8064Д | 1000 | |
| ППК-8122, ППК-8122М | 900 | |
| Контакты вспомогательной цепи: | ||
| Номинальный ток, А | 2 | |
| Номинальное напряжение, В | 110 | |
| Номинальное напряжение катушки вентиля | ||
| пневматического привода, В | ||
| ППК-8023, ППК-8023М | 24,75 | |
| ППК-8063 | 24 | |
| ППК-8063, ППК-8063М | 75 | |
| ППК-8122, ППК-8122М, ППК-8042, ППК-8042М, ППК-8064 | 110 | |
| Номинальное давление воздуха, МПа | 0,5 | |
| Максимальное давление воздуха, МПа | 0,675 | |
| Минимальное давление воздуха, МПа | 0,35 | |
| Масса, кг | ||
| ППК-8023, ППК-8023М | 100 | |
| ППК-8042, ППК-8042М | 132 | |
| ППК-8122, ППК-8122М, ППК-8063, ППК-8063М, ППК-8064, ППК-8064Д | 152 | |
| Климатическое исполнение | У2, Т3 | |
ППК12201 ППК12202 ППК12022 ППК12404Д ППК12044Д ППК 12602 ППК12062 ППК12602Д ППК12062Д ППК12802 ППК12082 ППК12804Д ППК12084Д ППК12801 ППК12081 ППК11201 ППК11602 ППК11062 | Контакты главной цепи: | |
| Номинальное напряжение для серии ППК 12000 | ||
| ППК11000, В | 1000 | |
| Род тока | постоянный | |
| Номинальный ток для серии ППК 12000, А | 1000 | |
| Номинальный ток для серии ППК 11000, А | 800 | |
| Контакты вспомогательной цепи: | ||
| Номинальный ток, А | 2 | |
| Номинальное напряжение катушки вентиля пневматического привода ивспомогательных цепей, В | ||
| Для серии ППК 12000, 11000, В | ||
| ППК 12201, 12801, 12081 | 75 | |
| ППК12202, 12022, 12404Д, 12044Д, 12602, 12062 | 110 | |
| 12602Д, ППК 12062Д, 12082, 12802, 12804Д, 12084Д, 11602, 11062 | ||
| Давление воздуха в приводе для серии переключателей ППК 12000, 11000: | ||
| Номинальное давление воздуха, МПа | 0,5 | |
| Максимальное давление воздуха, МПа | 0,675 | |
| Минимальное давление, МПа | 0,35 | |
| Масса, кг | ||
| ППК12404Д, 12044Д | 70 | |
| ППК 12201, 12202, 12022 | 50 | |
| ППК 12602, 12062, 12602Д, 12062Д | 95 | |
| ППК 12802,12082, 12804Д, 12084Д, 12801, 12081 | 105 | |
| ППК 11201 | 45 | |
| ППК 11602, 11062 | 87 | |
| Климатическое исполнение | У2, УХЛ3, Т3 | |
| ППК 12000, ППК 11000 |
Переключатель пневматический кулачковый типа ППК-8063У3 — FINDOUT.
 SU
SUПредставляет собой групповой коммутационный аппарат с электропневматическим приводом, обеспечивающим переключения в силовых цепях тепловоза при обесточенном состоянии этих цепей.
Конструкция аппарата включает корпус с приводом диафрагменного типа, вал управления с кулачковыми шайбами, главные и вспомогательные контактные группы.
Корпус переключателя является сборочной единицей, состоящей из стальных основания, кронштейна и шести вертикальных несущих стоек. Стойки выполнены шестигранными и связывают между собой основание и кронштейн. К боковым сторонам основания прилиты тарельчатые диски, к которым по фланцам прикрепляют диафрагмы с крышками привода переключателя. Образующиеся внутренние полости между диафрагмами и выпуклыми наружу крышками являются рабочими камерами привода. Диафрагмы выполнены из обрезиненной ткани и связаны друг с другом посредством штока, свободно пропущенного через отверстия, просверленные соосно в тарельчатых дисках. Крепление штока к диафрагмам производится через опорные шайбы.
В средней части к штоку болтами крепится накладка, имеющая по центру коническое отверстие. С помощью накладки шток соединяется с поводком вала управления. Для подачи сжатого воздуха в соответствующую рабочую камеру привода к основанию аппарата крепятся два электропневматических вентиля типа ВВ-32У3. Клапанные части вентилей связываются с рабочими камерами привода воздухопроводами.
Вал управления с кулачковыми шайбами обеспечивает переключение всех контактных групп аппарата. Он состоит из стального стержня, имеющего на длине участки круглого и квадратного сечений. На участок квадратного сечения вала устанавливаются кулачковые (профильные) пластмассовые шайбы в количестве шести штук. Профиль каждой шайбы позволяет переключать четыре пары главных контактов, а в случае отключения пневмопривода переключателя – удерживать эти контакты в конечном положении. Шайбы разнесены по длине участка с помощью распорных изолирующих втулок и закреплены корончатой гайкой под нижней шайбой. Верхняя шайба при этом упирается в стальное заштифтованное на валу кольцо с обоймой. На участке круглого сечения вала, выше шайб, через сквозное радиальное отверстие пропущен поводок – болт со сферической головкой. На резьбовой хвостовик поводка устанавливается скоба переключения вспомогательных контактных групп аппарата. Подшипниковыми узлами вала управления служат: по центру основания – втулка, закрепляемая болтами; по центру кронштейна – втулка, запрессованная в его расточку. Втулки имеют штуцерные маслёнки, заправляемые смазкой ЦИАТИМ-201.
Верхняя шайба при этом упирается в стальное заштифтованное на валу кольцо с обоймой. На участке круглого сечения вала, выше шайб, через сквозное радиальное отверстие пропущен поводок – болт со сферической головкой. На резьбовой хвостовик поводка устанавливается скоба переключения вспомогательных контактных групп аппарата. Подшипниковыми узлами вала управления служат: по центру основания – втулка, закрепляемая болтами; по центру кронштейна – втулка, запрессованная в его расточку. Втулки имеют штуцерные маслёнки, заправляемые смазкой ЦИАТИМ-201.
Главные контактные группы (24 пары контактов) смонтированы на шестигранных стойках переключателя: подвижные контакты – на центральных стойках, неподвижные – на угловых. Подвижные пальцевые контакты парами закреплены на качающихся рычагах и имеют гибкие медные выводы на изолированные контактодержатели.
Реверсор тепловоза ПК-8023 ТЭМ-2.
Служит для подключения к якорной обмотке тягового двигателя его обмотки возбуждения в положении «вперед» или «назад». На электрической схеме обозначается буквой «Р», контакты для подключения вперед в электрической схеме указаны замкнутыми, а для положения «назад» разомкнутыми.
На электрической схеме обозначается буквой «Р», контакты для подключения вперед в электрической схеме указаны замкнутыми, а для положения «назад» разомкнутыми.
В качестве реверсора применяют переключатель ППК-8023. Переключатель имеет корпус. Состоящий из двух панелей (верхней и нижней) и 5 шестигранных стоек. В обеих панелях имеются отверстия, в которые запрессованы подшипники качения. В них установлен вал переключателя.
Вал в средней части выполнен с квадратным сечением и на эту часть вала одет кулачковый текстолитовый барабан, с 4 кулачками, расположенные перпендикулярно друг другу. В верхней части вал имеет поводок пневмопривода и скобу переключения блок контактов. Силовые контакты располагаются на 3 передних стойках, к ним крепят изоляционные колодки. К колодкам крайних стоек крепят неподвижные силовые контакты, они попарно объединяются стальными планками. Планка оборудуется контактными выводами для крепления кабелей.
К изоляционной колодке средней стойки крепят медное ленточное соединение подвижных контактов
Сами подвижные контакты укрепляются на плечах, коромысла которых шарнирно укреплено на изоляционной колодке средней стойки. На концах коромысла пружины, коромысло, взаимодействуя с кулачками барабана может замыкать или размыкать соответствующие силовые контакты.
На концах коромысла пружины, коромысло, взаимодействуя с кулачками барабана может замыкать или размыкать соответствующие силовые контакты.
Пневмо привод выполнен из корпуса, отлит за одно с верхней панелью. По торцам корпус с вогнутыми фланцами, к которым крепятся выпуклые крышки. Между фланцем и крышкой находится диафрагма. Диафрагма состоит из стальной шайбы с привулканизированной к ней резиной, на его верхнем конце крепят шестигранную головку под ключ.
При дистанционном управлении подается питание на катушку одного из электропневматических вентиля, укрепленных на верхней панели. Он открывает доступ воздуха к соответствующей диафрагме и она, прогибаясь, перемещает шток в нужную сторону, и он воздействует на поводок поворачивает вал. Шайба укрепляется на концах штока. В средней части к штоку крепится две пластины, между торцами которых образуется зазор, в который входит поводок вала реверсора. На верхней панели крепят две группы блок контактов траверсно мостикового типа. Траверсы этих групп могут входить в взаимодействие со скобой вала реверсора. Вал реверсора может поворачиваться либо с помощью пневмо привода либо в ручную.
Траверсы этих групп могут входить в взаимодействие со скобой вала реверсора. Вал реверсора может поворачиваться либо с помощью пневмо привода либо в ручную.
ООО ЖелПромСнаб г. Красноярск, новости
Возбудитель ВС-650 27.08.2014 01:21В наличии на складе возбудитель ВС-650. Новый. Производство Украина. 2010 год.
Гильзы Д100 03.07.2014 13:45
На склад поступили Гильзы цилиндра Д100.01.101. 20 штук.
Поршни Д49 11.05.2014 18:18
В наличии поршни дизеля Д49 различных исполнений.
Поршень 1-5Д49.22 спч
Поршень 2-5Д49.22 спч
Поршень 4-5Д49.22.1 спч02
Поршень 4-5Д49.22.1 спч04
Поршень 5Д49. 22 спч 9-01
22 спч 9-01
Производство Пензадизельмаш и Коломенский завод.
А так же головки и тронки отдельно.
Вкладыши Д100 30.03.2014 21:14
Большое поступление вкладышей дизеля Д100. Коренные, шатунные, разные градации!
Кольца поршневые КТ-6 13.03.2014 04:28
В наличии на складе запасные части к компрессору КТ6. Поршни, цилиндры, кольца поршневые разных градаций (0,1,2,3,4).
Полюс генератора ПМ111 ОМ-5 22.12.2013 19:41
В наличии на складе полюса для генератора постоянного тока ПМ111-ОМ5.
Звоните 8-902-962-4042 Юрий.
Контакторы, реле, датчики 17.12.2013 18:36
В наличии на складе Контакторы ПК1146 110В, датчики температуры ТАМ103С, реле ТРПУ6-2-110В-П и другое электрооборудование!
Крышки цилиндра Д50 17.
 11.2013 19:23
11.2013 19:23В наличии крышки цилиндра Д50! В сборе с клапанами и без клапанов.
| Наименование | Цена за ед. | Кол-во |
Ед. изм. изм.
|
Сумма |
|---|---|---|---|---|
| Контакт 3Б-02150 | не указана | 15 | Штука | не указана |
Контакт 5ТЕ. 551.009 551.009
|
не указана | 5 | Штука | не указана |
| Контакт 3Б-02151 | не указана | 15 | Штука | не указана |
Контакт 5ТЛ. 551.272 551.272
|
не указана | 10 | Штука | не указана |
Контакт 5ТЛ. 551.271 551.271
|
не указана | 10 | Штука | не указана |
Контакт 5ТХ.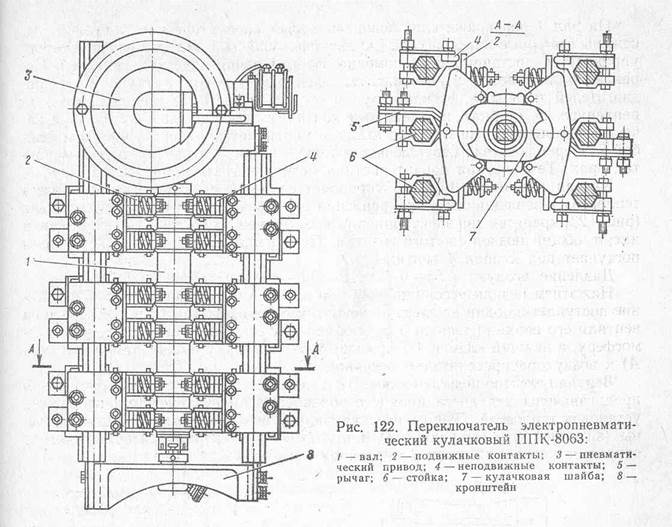 551.100 ППК-8063 551.100 ППК-8063
|
не указана | 10 | Штука | не указана |
Контакт 5ТН.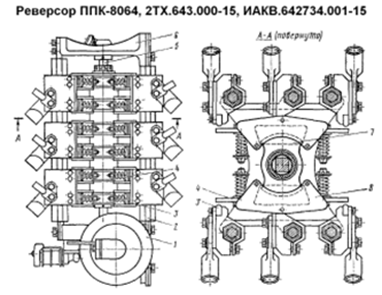 551.209 551.209
|
не указана | 80 | Штука | не указана |
Контакт 5ТЛ. 551.193 RA-221 49Х10Х3 подвижный на ру 551.193 RA-221 49Х10Х3 подвижный на ру
|
не указана | 150 | Штука | не указана |
Контакт 5ТХ. 551.100-01 551.100-01
|
не указана | 10 | Штука | не указана |
Контакт 8ТЕ. 551.057 551.057
|
не указана | 20 | Штука | не указана |
Контакт 5ТХ. 551.199 551.199
|
не указана | 10 | Штука | не указана |
Контакт 5ТХ. 551.195 неподвижный 551.195 неподвижный
|
не указана | 10 | Штука | не указана |
Контакт 5ТХ. 551.189 БИЛТ.685179.040 подвижный 551.189 БИЛТ.685179.040 подвижный
|
не указана | 6 | Штука | не указана |
Контакт 8ТЕ. 551.085 551.085
|
не указана | 20 | Штука | не указана |
Контакт 5ТХ. 551.199-01 подвижный 551.199-01 подвижный
|
не указана | 10 | Штука | не указана |
Контакт 8ТЛ. 551.037 (8ТН.551.023, ПК-31 ЭТ.063) 551.037 (8ТН.551.023, ПК-31 ЭТ.063)
|
не указана | 3105 | Штука | не указана |
Контакт 8ТЛ. 551.037 8ТН.551.023 ЭТ.063 41Х34Х25 551.037 8ТН.551.023 ЭТ.063 41Х34Х25
|
не указана | 100 | Штука | не указана |
Контакт 8ТЛ. 551.003 8ТЕ.551.008 БВП502 175Х60Х65 неподвижный 551.003 8ТЕ.551.008 БВП502 175Х60Х65 неподвижный
|
не указана | 14 | Штука | не указана |
Контакт 8ТЛ. 551.006 (8ТЕ.551.009) БВП-5-02 82Х44Х33 подвижн 551.006 (8ТЕ.551.009) БВП-5-02 82Х44Х33 подвижн
|
не указана | 22 | Штука | не указана |
Контакт 8ТЛ. 551.074 (для 1КМ) подвижный 551.074 (для 1КМ) подвижный
|
не указана | 34 | Штука | не указана |
Контакт 8ТЛ. 551.012 неподвижный 551.012 неподвижный
|
не указана | 15 | Штука | не указана |
Контакт 8ТЛ. 551.028 (8ТН.551.045) ПК-14-19 41Х34Х20 551.028 (8ТН.551.045) ПК-14-19 41Х34Х20
|
не указана | 80 | Штука | не указана |
Контакт 8ТХ.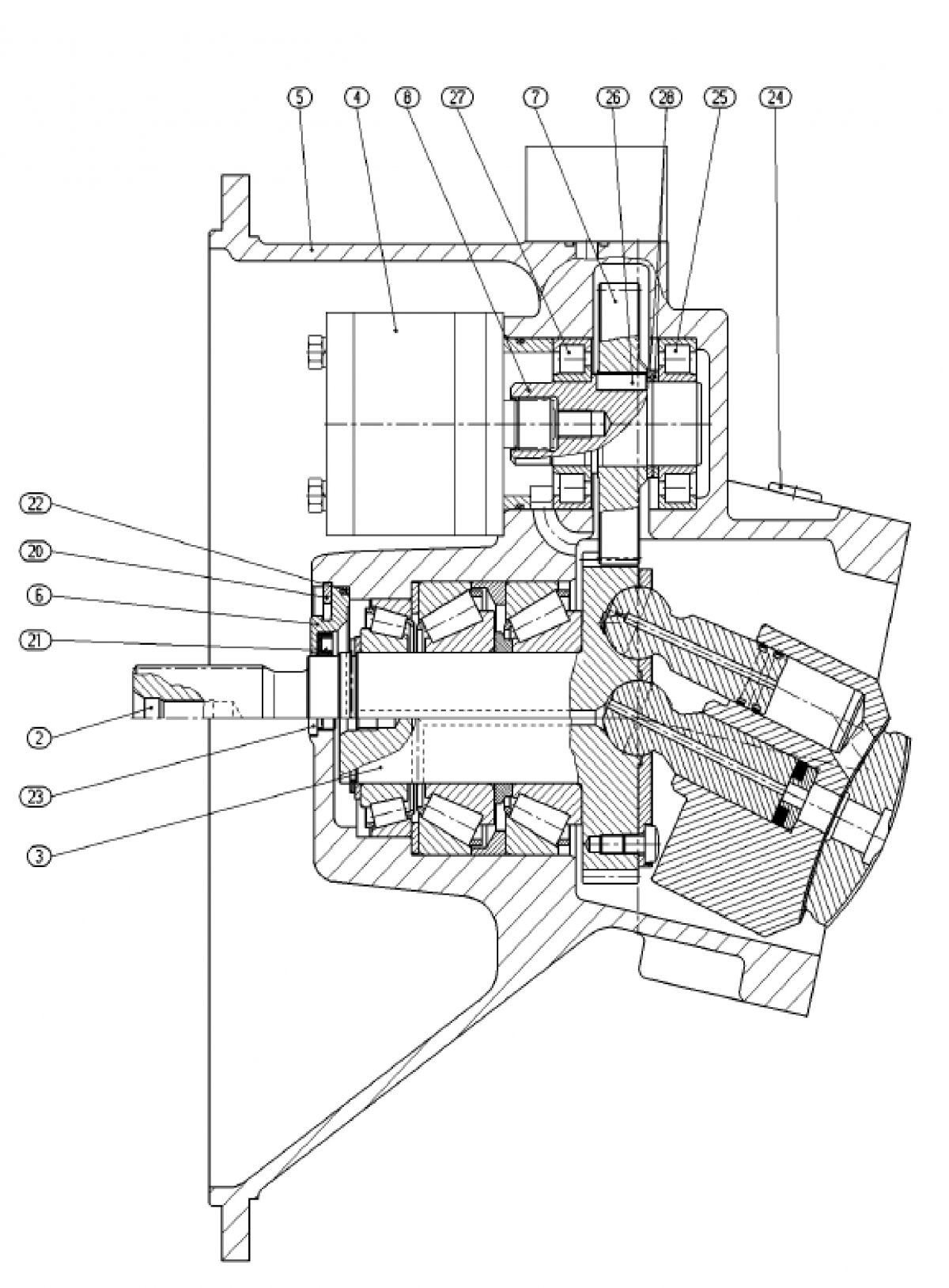 551.058 силовой 551.058 силовой
|
не указана | 20 | Штука | не указана |
Контакт 8ТН. 551.078 неподвижный 551.078 неподвижный
|
не указана | 8 | Штука | не указана |
Контакт 8ТХ. 551.072 551.072
|
не указана | 20 | Штука | не указана |
Контакт АК-11Б ТИБЛ. 685114.002 подвижный 685114.002 подвижный
|
не указана | 115 | Штука | не указана |
Контакт АКБ-11Б ТИБЛ. 685114.001 неподвижный 685114.001 неподвижный
|
не указана | 85 | Штука | не указана |
Контакт МК-310Б МК-204 8ТН. 551.125 электромагнит контактора 551.125 электромагнит контактора
|
не указана | 45 | Штука | не указана |
Контакт 8ТН. 551.077 неподвижный 551.077 неподвижный
|
не указана | 8 | Штука | не указана |
Контакт неподвижный 5ЛХ. 143.247 143.247
|
не указана | 6 | Штука | не указана |
Контакт МК-6 ГЛЦИ. 685161.113 25Х73Х3 подвижный контактора 685161.113 25Х73Х3 подвижный контактора
|
не указана | 100 | Штука | не указана |
Контакт неподвижный БК-78 ЭТ. 030 030
|
не указана | 20 | Штука | не указана |
| Контакт МК-6 неподвижный | не указана | 200 | Штука | не указана |
Контакт неподвижный БИЛТ. 685179.033 685179.033
|
не указана | 6 | Штука | не указана |
Контакт подвижный БКЖИ. 685172.004 685172.004
|
не указана | 6 | Штука | не указана |
Контакт Т93. 81.14.00 подвижный 81.14.00 подвижный
|
не указана | 200 | Штука | не указана |
Соединение 5ТЛ. 505.004 гибкое 505.004 гибкое
|
не указана | 20 | Штука | не указана |
Соединение гибкое 5ТЛ. 505.000 (для контактора 1КМ.016М) 505.000 (для контактора 1КМ.016М)
|
не указана | 20 | Штука | не указана |
Планка БИЛТ. 741.612.015 ПК-1146А 75В 741.612.015 ПК-1146А 75В
|
не указана | 76 | Штука | не указана |
Мостик ПК-1616А У3 БИЛТ. 685171.007 685171.007
|
не указана | 2 | Штука | не указана |
Группа контактная БИЛТ. 685162.001 685162.001
|
не указана | 30 | Штука | не указана |
Alcatel 8063.
Как установить официальную стоковую прошивку на Alcatel PIXI 4 (5045x)Мы рекомендуем использовать последнюю версию прошивки tct alcatel pixi4 7 wifi. Простое пошаговое обновление прошивки до последней версии, скачивание когда-либо выпускается. Загрузите и обновите прошивку для моделей: tecno camon 15 pro cd8, tecno camon 15 premier cd8j, tct alcatel 3x plus tokyopro a, oppo a52 op4c7bl1 cph, blu studio m4 plus su. Все права защищены. Несанкционированное воспроизведение полностью или частично строго запрещено.Оливия:. Сэди:. Харман :.
Я только что загрузил это приложение. Бетти:. Франни :. Набор:. Альберт :. Ванесса :. Киа :. Это не всегда срабатывает при попытке восстановить историю. Мне было очень комфортно со старой версией. Лекси :. Загрузите прошивку Samsung Galaxy S5, почти каждый сегодня знает, что такое операционная система Android, почему она так популярна среди миллионов пользователей и как использовать весь ее потенциал, как получить root права на мобильный телефон Samsung Galaxy Android и выкорчевать его, используя стандартную прошивку или ПЗУ.
Существует множество различных мобильных прошивок и настраиваемых ПЗУ для рутированных устройств Android. Но иногда приходится возвращаться к стоковой прошивке. Скачать гаджет для обновления прошивки ZTE очень важно, так как многие его владельцы регулярно делают обновления, чтобы их гаджеты работали лучше. Обновление прошивки ZTE Stock ROM Flash File — это политика компании, чтобы клиенты, решившие оставить себе телефоны или планшеты на долгое время, могли пользоваться его лучшими возможностями.
ССЫЛКА В этой статье будет объяснено, зачем это нужно, как проверить обновление, а также как его загрузить и установить.Как загрузить прошивку Vodafone, несомненно, является крупнейшим поставщиком мобильных телефонов и интернет-услуг, поэтому любой человек, который становится клиентом компании, может задаться вопросом, как обновить прошивку для Vodafone. ССЫЛКА Обычно люди могут быть заинтересованы и нуждаться в загрузке и установке прошивки для модема, маршрутизатора, широкополосного доступа или некоторых телефонов, которые производятся крупными производителями специально для Vodafone и содержат только название компании.
Здесь мы рассмотрим, как делать это эффективно и безопасно.Скачивание и обновление прошивки Прошивка Coolpad производит очень популярные мобильные телефоны и планшеты, которые чрезвычайно популярны во многих странах по всему миру, поэтому обновления прошивки Coolpad могут быть полезны каждому владельцу устройств.
Как и у любого другого производителя. LINK Coolpad старается обновлять свои продукты, поэтому, даже если вы являетесь владельцем телефона в течение нескольких лет, его все равно можно модернизировать, чтобы повысить его эффективность и возможности. В этой статье мы постараемся объяснить, как проверить, скачать и установить прошивку Coolpad.Обновление прошивки LG может потребоваться всем, кто пользуется гаджетами и продуктами компании в течение длительного периода времени. Компания много делает для обслуживания клиентов и прилагает максимум усилий, чтобы ее продукция оставалась неизменной в соответствии с текущими требованиями рынка, несмотря на то, что она была выпущена задолго до этого.
LINK Клиенты всегда могут быть уверены, что возможности гаджетов можно расширить, загрузив и установив обновленную прошивку, а в этой статье мы объясним, как это сделать.Как установить и мы рекомендуем использовать последнюю версию прошивки tct alcatel pixi4 7 wifi.
108 muniswaran names на тамильскомПростое пошаговое обновление прошивки последней версии, возможность скачивания любого выпуска. Загрузите и обновите прошивку для моделей: tecno camon 15 pro cd8, tecno camon 15 premier cd8j, tct alcatel 3x plus tokyopro a, oppo a52 op4c7bl1 cph, blu studio m4 plus su. Как загрузить и обновить прошивку Прошивка tct alcatel pixi4 7 wifi Скачать прошивку для Samsung Galaxy S5 Сегодня почти каждый знает, что такое операционная система Android, почему она так популярна среди миллионов пользователей и как использовать весь ее потенциал, как получить root права на Android Samsung Galaxy мобильный телефон и отключите его, используя стоковую прошивку или ПЗУ.
Существует множество различных мобильных прошивок и настраиваемых ПЗУ для рутированных устройств Android. Но иногда приходится возвращаться к стоковой прошивке. Скачать гаджет для обновления прошивки ZTE очень важно, так как многие его владельцы регулярно делают обновления, чтобы их гаджеты работали лучше. Обновление прошивки ZTE Stock ROM Flash File — это политика компании, чтобы клиенты, решившие оставить себе телефоны или планшеты на долгое время, могли пользоваться его лучшими возможностями.
Файл прошивки Alcatel Tab 8063 Mt8127 6.0 Marshmallow Скачать
ССЫЛКА В этой статье будет объяснено, зачем это нужно, как проверить обновление, а также как его загрузить и установить. Как загрузить прошивку Vodafone, несомненно, является крупнейшим поставщиком мобильных телефонов и интернет-услуг, поэтому любой человек, который становится клиентом компании, может задаться вопросом, как обновить прошивку для Vodafone. ССЫЛКА Обычно люди могут быть заинтересованы и нуждаться в загрузке и установке прошивки для модема, маршрутизатора, широкополосного доступа или некоторых телефонов, которые производятся крупными производителями специально для Vodafone и содержат только название компании.
Здесь мы рассмотрим, как делать это эффективно и безопасно. Скачивание и обновление прошивки Прошивка Coolpad производит очень популярные мобильные телефоны и планшеты, которые чрезвычайно популярны во многих странах по всему миру, поэтому обновления прошивки Coolpad могут быть полезны каждому владельцу устройств.
Как и у любого другого производителя. LINK Coolpad старается обновлять свои продукты, поэтому, даже если вы являетесь владельцем телефона в течение нескольких лет, его все равно можно модернизировать, чтобы повысить его эффективность и возможности.В этой статье мы постараемся объяснить, как проверить, скачать и установить прошивку Coolpad. Обновление прошивки LG может потребоваться всем, кто пользуется гаджетами и продуктами компании в течение длительного периода времени.
Компания много делает для обслуживания клиентов и прилагает максимум усилий, чтобы ее продукция оставалась неизменной в соответствии с текущими требованиями рынка, несмотря на то, что она была выпущена задолго до этого. ССЫЛКА Клиенты всегда могут быть уверены, что возможности гаджетов можно расширить, загрузив и установив обновленную прошивку, а в этой статье мы объясним, как это сделать.
Как установить и добро пожаловать на форумы GSM-Forum. В настоящее время вы просматриваете наши форумы в качестве гостя, что дает вам ограниченный доступ к просмотру большинства обсуждений и другим нашим функциям. Только зарегистрированные участники могут задавать вопросы, связываться с другими участниками или выполнять поиск в нашей базе данных, содержащей более 8 миллионов сообщений. Если у вас возникли проблемы с регистрацией или входом в учетную запись, свяжитесь с нами.
Сообщений: 2, Участник: Статус: Offline. Последний раз редактировалось Brka; Причина: объединенные посты.Сообщений: 6, Участник: Статус: Offline Я нашел флеш-файл для вашего планшета. Зацените завтра, я выложу его здесь. Ваше устройство отображается как порт MTK в диспетчере устройств?
Утвердить расширение ядра Mac терминалСообщений: 2, Член: Статус: Offline привет не указан как порт MTK Сообщений: 6, Член: Статус: Offline Цитата :. Сообщение от zubairhussain. Сообщений: 3 Участник: Статус: Offline Мне нужна прошивка или файл образа для alcatel pixi4 7-дюймовая модель планшета с Wi-Fi зефир Vers 6. Первоначально Послал algomac.Сообщений: 30 Участник: Статус: Offline привет, мне нужна прошивка или файл образа для alcatel pixi4 7-дюймовая модель планшета с Wi-Fi, спасибо.
Сообщений: 2, Участник: Статус: Offline Цитата :.
Нельсон Сэсслер возрастСообщение от Brka. Нашел флешку для вашего планшета Сообщение от ahmedksal. Первоначально Послано ghandour Если вы разместите оригинальный PID, и прошивка доступна, возможно, я смогу помочь. Причина: обновленная ссылка. Вот и причина: удалена ссылка. Сообщений: 2 Участник: Статус: Offline Помогите, пожалуйста, как сделать это устройство root.Первоначально Послано heartache Помогите, пожалуйста, как сделать это устройство root.
BB код включен. Смайлы включены. Трекбэки включены. Мы рекомендуем использовать последнюю версию прошивки tct alcatel pixi4 7 wifi устройства. Простое пошаговое обновление прошивки до последней версии, скачивание когда-либо выпускается. Загрузите и обновите прошивку для моделей: tecno camon 15 pro cd8, tecno camon 15 premier cd8j, tct alcatel 3x plus tokyopro a, oppo a52 op4c7bl1 cph, blu studio m4 plus su. Все права защищены.
Несанкционированное воспроизведение, полное или частичное, строго запрещено.Манда :. Только начал пользоваться приложением. Глория :. Алина :. Кендалл :. Nervie :. Мелли :.
Alcatel Pixi 4 7 ″ WiFi OT 8063 Флэш-файл со стоковой прошивкой
Рональд :. Лансон :. Захария :. Мэдисон :. Мне нравятся твои приложения! Вы великолепны. Приложения всегда делают меня и моих братьев и сестер счастливее — мое любимое приложение !!
Загрузите прошивку Samsung Galaxy S5. Сегодня почти каждый знает, что такое операционная система Android, почему она так популярна среди миллионов пользователей и как использовать весь ее потенциал, как получить root права на мобильный телефон Samsung Galaxy Android и выкорчевать его, используя стандартную прошивку или ПЗУ. .Существует множество различных мобильных прошивок и настраиваемых ПЗУ для рутированных устройств Android. Но иногда приходится возвращаться к стоковой прошивке. Скачать гаджет для обновления прошивки ZTE очень важно, так как многие его владельцы регулярно делают обновления, чтобы их гаджеты работали лучше.
Обновление прошивки ZTE Stock ROM Flash File — это политика компании, чтобы клиенты, решившие оставить телефоны или планшеты на долгое время, могли пользоваться его лучшими возможностями.
ССЫЛКА В этой статье будет объяснено, зачем это нужно, как проверить обновление, а также как его загрузить и установить.Как загрузить прошивку Vodafone, несомненно, является крупнейшим поставщиком мобильных телефонов и интернет-услуг, поэтому любой человек, который становится клиентом компании, может задаться вопросом, как обновить прошивку для Vodafone.
ССЫЛКА Обычно люди могут быть заинтересованы и нуждаться в загрузке и установке прошивки для модема, маршрутизатора, широкополосного доступа или некоторых телефонов, которые производятся крупными производителями специально для Vodafone и содержат только название компании.
Alcatel Pixi 4 7 ″ WiFi OT 8063 Флэш-файл стандартной прошивки
Здесь мы рассмотрим, как сделать это эффективно и безопасно.Скачивание и обновление прошивки Прошивка Coolpad производит очень популярные мобильные телефоны и планшеты, которые чрезвычайно популярны во многих странах по всему миру, поэтому обновления прошивки Coolpad могут быть полезны каждому владельцу устройств.
Как и у любого другого производителя. LINK Coolpad старается обновлять свои продукты, поэтому, даже если вы являетесь владельцем телефона в течение нескольких лет, его все равно можно модернизировать, чтобы повысить его эффективность и возможности. В этой статье мы постараемся объяснить, как проверить, скачать и установить прошивку Coolpad.Обновление прошивки LG может потребоваться всем, кто пользуется гаджетами и продуктами компании в течение длительного периода времени. Компания много делает для обслуживания клиентов и прилагает максимум усилий, чтобы ее продукция оставалась неизменной в соответствии с текущими требованиями рынка, несмотря на то, что она была выпущена задолго до этого.
LINK Клиенты всегда могут быть уверены, что возможности гаджетов можно расширить, загрузив и установив обновленную прошивку, а в этой статье мы объясним, как это сделать.Как установить и прошивку Samsung: будучи одним из ведущих производителей электронных устройств, в том числе мобильных телефонов, Samsung очень часто выпускает Flash-файл прошивки прошивки для своих продуктов. У большого количества людей могут возникнуть вопросы о том, как загрузить обновление прошивки Samsung для мобильных устройств, поэтому мы постараемся объяснить это ниже.
Процедура довольно проста и в большинстве случаев успех гарантирован даже не опытным пользователям. Мы рекомендуем использовать последнюю версию прошивки tct alcatel pixi4 7 wifi.Простое пошаговое обновление прошивки до последней версии, скачивание когда-либо выпускается.
Загрузите и обновите прошивку для моделей: tecno camon 15 pro cd8, tecno camon 15 premier cd8j, tct alcatel 3x plus tokyopro a, oppo a52 op4c7bl1 cph, blu studio m4 plus su. Все права защищены. Несанкционированное воспроизведение полностью или частично строго запрещено. Августина :. Криста :. Кармон :. Мюриэль :. Рениус :. Это 2-е изменение в приложении.
Не нравится мне это обновление. Старый пользоваться было проще.Может:. Стержень:. Дженни:.
Мне нравятся ваши приложения! Вы великолепны. Приложения всегда делают меня и моих братьев и сестер счастливее — мое любимое приложение !! Дилли :. Харти :. Это одно из лучших приложений на свете! Загрузите прошивку Samsung Galaxy S5, почти каждый сегодня знает, что такое операционная система Android, почему она так популярна среди миллионов пользователей и как использовать весь ее потенциал, как получить root права на мобильный телефон Samsung Galaxy Android и выкорчевать его, используя стандартную прошивку или ПЗУ.
Существует множество различных мобильных прошивок и настраиваемых ПЗУ для рутированных устройств Android.Но иногда приходится возвращаться к стоковой прошивке. Скачать гаджет для обновления прошивки ZTE очень важно, так как многие его владельцы регулярно делают обновления, чтобы их гаджеты работали лучше. Схема подключения пропановой печи
Базовая схема подключения на веб-сайтеОбновление прошивки ZTE Stock ROM Flash File — это политика компании, чтобы клиенты, которые решили оставить телефоны или планшеты на долгое время, могли пользоваться его лучшими возможностями. ССЫЛКА В этой статье будет объяснено, зачем это нужно, как проверить обновление, а также как его загрузить и установить.
Как загрузить прошивку Vodafone, несомненно, является крупнейшим поставщиком мобильных телефонов и интернет-услуг, поэтому любой человек, который становится клиентом компании, может задаться вопросом, как обновить прошивку для Vodafone. ССЫЛКА Обычно люди могут быть заинтересованы и нуждаться в загрузке и установке прошивки для модема, маршрутизатора, широкополосного доступа или некоторых телефонов, которые производятся крупными производителями специально для Vodafone и содержат только название компании.
Здесь мы рассмотрим, как делать это эффективно и безопасно.Скачивание и обновление прошивки Прошивка Coolpad производит очень популярные мобильные телефоны и планшеты, которые чрезвычайно популярны во многих странах по всему миру, поэтому обновления прошивки Coolpad могут быть полезны каждому владельцу устройств.
Как и у любого другого производителя. LINK Coolpad старается обновлять свои продукты, поэтому, даже если вы являетесь владельцем телефона в течение нескольких лет, его все равно можно модернизировать, чтобы повысить его эффективность и возможности. В этой статье мы постараемся объяснить, как проверить, скачать и установить прошивку Coolpad.Обновление прошивки LG может потребоваться всем, кто пользуется гаджетами и продуктами компании в течение длительного периода времени.
Компания много делает для обслуживания клиентов и прилагает максимум усилий, чтобы ее продукция оставалась неизменной в соответствии с текущими требованиями рынка, несмотря на то, что она была выпущена задолго до этого. ССЫЛКА Клиенты всегда могут быть уверены, что возможности гаджетов можно расширить, загрузив и установив обновленную прошивку, а в этой статье мы объясним, как это сделать.Как установить и Этот пакет прошивки состоит из трех разных файлов образа: boot. Причина, по которой эта прошивка загружается отдельно по разделам, состоит в том, чтобы упростить для тех, кому просто нужно прошить только один раздел, чтобы исправить свое заблокированное устройство или по другим связанным причинам.
После загрузки файлов прошивки s для прошивки на Pixi 7 по ссылкам выше, следуйте простому руководству ниже, чтобы прошить изображения. Как только будет запущена команда перезагрузки, ваше устройство автоматически перезагрузится в систему.Первая загрузка обычно выполняется медленно, в то время как на некоторых устройствах это похоже на перезагрузку устройства. Так что наберитесь терпения и дождитесь полной загрузки устройства. Сообщите мне о последующих комментариях по электронной почте. Уведомлять меня о новых сообщениях по электронной почте. Этот сайт использует Akismet для уменьшения количества спама.
Узнайте, как обрабатываются данные вашего комментария.
Alcatel Pixi 4 8063 MT8127 6.0 Обход FRP Cuenta GoogleПохожие сообщения. Коллинз Эдим 21 марта, Все ссылки исправлены и обновлены. Спасибо за внимание! Люк 14 августа, Alcatel one touch pixi 3 7х прошивка пожалуйста.Коллинз Эдим 30 ноября, Андриус Осворт 29 ноября, Аноним 25 сентября, Оставить комментарий Отменить ответ Сообщите мне о последующих комментариях по электронной почте. Помогите остановить Коронавирус! Оставайтесь дома, если возможно, и оставайтесь в безопасности! XDA Developers была основана разработчиками для разработчиков.
Теперь это ценный ресурс для людей, которые хотят максимально использовать свои мобильные устройства, от настройки внешнего вида до добавления новых функций. Вы разработчик? Условия использования. Размещено на Leaseweb. LineageOS ColorOS 7.Обзор Realme 6 Pro: всесторонний доступный смартфон с несколькими премиальными функциями 29 марта, счетчик благодарностей: 1, предупреждение: текущий Pixi 4 4 N будет мягким, если вы разблокируете вспышку.
OP Неактивный признанный участник. Дата регистрации: Присоединился: октябрь. Сейчас я размещаю информацию о различных моделях устройств Pixi 4 и связываю их с соответствующими публикациями. Со временем это будет редактироваться, так что следите за обновлениями. Как только я получу способ сделать резервную копию и смогу восстановить эту резервную копию, я добавлю ее сюда.Alcatel OT Pixi Должен уметь работать на Pixi 4 4 «только меняем рекавери. Порт MT 6.
ПЗУ Pixi 4. Требуются базовые навыки работы с компьютером, а также минимальные знания о телефоне и телефонных утилитах. Убедитесь, что все, что вы делаете, вы делаете под свою ответственность.
Счетчик благодарностей: Дата регистрации: Присоединился: июн Привет, спасибо за подписку и оставьте свой отзыв. Полагаю, ты говоришь, что это не сработало?
Вы сначала сделали резервную копию ПЗУ, можете ли вы нам ее предоставить? Если нет, можете ли вы вернуться и сделать это? Счетчик благодарностей: 1.
Google Play вылетел. KingRoot рутирован. Сначала Системное приложение Managerapp от flaki потом рухнуло. Они интегрированы в обновления.
Удалены Google Фото, Google Книги и тому подобное. Но никаких системных файлов. После восстановления мобильного телефона. Wi-Fi соединение. Или подключение 3G.
| Вы можете летать коммерчески без лицензии пилота дрона, но вы должны зарегистрировать свой дрон и получить аккредитацию оператора RPA от CASA.Это бесплатно и удобный вариант для новичков, которые хотят просто запустить свой дрон на законных основаниях и начать зарабатывать деньги.
Помните, что полет на дроне для заработка на самом деле незаконен в Австралии, если у вас нет хотя бы аккредитации оператора RPA от CASA (или RePL или ReOC).
Вот 5 веских причин убедиться, что вы получили одобрение CASA перед полетом. Как получить аккредитацию оператора RPA от CASA: Вот 2 шага, которые вам нужно сделать, чтобы получить разрешение (от CASA) на управление дроном для коммерческого использования без лицензии: Шаг 1: зарегистрируйте свой дрон здесь, иначе вам грозит штраф до 11 100 долларов. | Если ваш дрон весит более 2 кг, и вы обычно предпочитаете летать для чужого бизнеса по производству дронов (а не начинаете свой бизнес), лучше всего получить лицензию удаленного пилота. Безусловно, самый популярный вариант лицензии. RePL обычно представляет собой 5-дневный курс, охватывающий теорию и практику, или теоретическую часть онлайн + 2 дня практической работы. Срок действия этой лицензии пилота дрона не истекает, и ее продление не требуется! Минимальный возраст — 17 лет. Получить курс | Лучше всего, если ваш дрон весит более 2 кг, и вы хотите создать собственный бизнес по производству дронов и работать напрямую с клиентами. Обычно для начинающих владельцев дронов, которые планируют нанимать отдельных пилотов и вести бизнес на полную ставку. Получение ReOC — это более сложный процесс, который требует, чтобы вы сначала получили RePL, а затем отправили процедуру и политику вашей компании в Управление безопасности гражданской авиации (CASA) для рассмотрения и утверждения. Наградой за получение этой сертификации является наличие большего количества возможностей и меньшее количество ограничений в отношении зон полетов, а также большее признание отрасли со стороны потенциальных клиентов, которые могут решить, что они хотят нанимать только пилотов с наивысшим доступным уровнем сертификации (держатели ReOC). Получить курс |
SMF100AT1G datasheet — Технические характеристики: Упаковка / ящик: SOD-123F; Упаковка:
IP4777CZ38 : DIODE, ESD, DVI HDMI INTERFACE, SOT510-1. s: Тип диода: Защита от электростатического разряда; Максимальное напряжение зажима Vc: 8 В; Рабочее напряжение: 5 В; Тип корпуса диода: -; Количество контактов: 38; Рассеиваемая мощность Pd: 14 мВт; MSL: -.
PESD12VS1UA : ДИОД, ESD, НИЗКАЯ ЕМКОСТЬ, SOD323. s: Тип диода: Защита от электростатического разряда; Максимальное напряжение зажима Vc: 27 В; Рабочее напряжение: — ; Тип корпуса диода: SOD-323; Нет.контактов: 2; Рассеиваемая мощность Pd: 360 мВт; MSL: -.
MCASP-L-E 10A : ПРЕДОХРАНИТЕЛЬ, АВТОМОБИЛЬНЫЙ, МИНИ-ЛЕЗВИЕ, 10 А, 32 В. s: Номинальное напряжение переменного тока: -; Номинальное напряжение постоянного тока: 32 В; Ток предохранителя: 10А; Размер предохранителя с автоматическим лезвием: 11,2 мм x 4 мм x 9 мм; Характеристики удара: -; Серия: MINI.
V9MLA0603NH : Варисторы 9V 20A 120pF. s: Производитель: Littelfuse; Категория продукта: Варисторы; RoHS: подробности; Продукт: MLV; Номинальное напряжение постоянного тока: 9 В; Емкость: 490 пФ; Диапазон рабочих температур: от -55 C до + 125 C; Монтаж: SMD / SMT; Упаковка: Катушка; Размеры: 0.8 мм Ш x 1,6 мм Д x 1 мм В; Упаковка / ящик: 0603 (1608 метрическая система); Количество заводской упаковки :.
046501.6DR : Предохранители 250V 1.6A Time Lag — UMF. Littelfuse предлагает широкий ассортимент предохранителей NANO ® с различными наборами функций для различных применений. Линия продуктов NANO Fuse включает более 15 серий высокопроизводительных устройств, в том числе Littelfuse серии 448 NANO²® SMF Fuse s. Предохранители серии 448 очень быстродействующие и имеют широкий.
ERB-SD2R50U : Предохранители Micro Chip Fuse0402.s: Производитель: Panasonic; RoHS: подробности; Текущий рейтинг: 2,5 А; Номинальное напряжение: 24 Вольт; Размер / группа предохранителей: 1005; Тип завершения: SMD / SMT; Тип предохранителя: быстродействующий; Материал корпуса: керамика; Размеры: 1 мм Д x 0,5 мм Ш; Упаковка: Катушка; Продукт: предохранитель Micro Chip; Рейтинг прерывания: 35 ампер; Монтаж.
170M5762 : Предохранитель — электрический, защита специальных цепей 630A, 690VAC, быстродействующий; ФГУП 630А 690В 2ФКЭ / 115 АР УК. с: Ток: 630А; Напряжение — номинальное: 690 В переменного тока; Упаковка / ящик: крепление на болтах; Тип предохранителя: быстродействующий; Тип установки: на болтах; Статус без содержания свинца: без содержания свинца; Статус RoHS: Соответствует RoHS.
0034.5624.22 : Защита цепи предохранителем 5A 250VAC, медленное срабатывание, задержка по времени; ПРЕДОХРАНИТЕЛЬ 5А 250В T-LAG 5X20 SMD T / R. с: Ток: 5А; Напряжение — номинальное: 250 В переменного тока; Упаковка / ящик: цилиндрический, 5×20 мм; Тип предохранителя: медленный, с задержкой по времени; Тип установки: держатель / поверхностное крепление; Статус без содержания свинца: без содержания свинца; Статус RoHS: Соответствует RoHS.
FWK-5A20F : Предохранитель — электрический, защита специальных цепей 5 А, 750 В переменного тока, 750 В постоянного тока, быстродействующий; ПРЕДОХРАНИТЕЛЬ 5A 750V 20X127MM FERRULE. с: Ток: 5А; Напряжение — номинальное: 750 В переменного тока, 750 В постоянного тока; Упаковка / ящик: цилиндрический; Тип предохранителя: быстродействующий; Тип установки: Держатель; Статус без содержания свинца: без содержания свинца; Статус RoHS: Соответствует RoHS.
0BLF010.T : Предохранитель — электрический, защита специальных цепей 10A 250VAC, быстродействующий; ПРЕДОХРАНИТЕЛЬ FAST 10A 250V MIDGET. s: Ток: 10А; Напряжение — номинальное: 250 В переменного тока; Корпус / Корпус: Цилиндрический, 5AG, 10,3×38,1 мм; Тип предохранителя: быстродействующий; Тип установки: Держатель; Статус без содержания свинца: без содержания свинца; Статус RoHS: Соответствует RoHS.
V510LA80BPX10 : ТВ — варисторы, защита цепи Mov 6.5kA 860V; РАДИАЛЬНЫЙ ВАРИСТОР 20ММ. s: Максимальное напряжение переменного тока: 510 В переменного тока; Максимальное напряжение постоянного тока: 675 В постоянного тока; Скачок тока: 6.5кА; Упаковка / футляр: диск 20 мм; Напряжение варистора: 860 В; Энергия: 190Дж; Количество цепей: 1; Статус без содержания свинца: без содержания свинца; Статус RoHS: Соответствует RoHS.
P0641CA2LRP : Телевизор — Другая составная защита цепи 58В -; СИДАКТОР 58В 90А НАПРЯЖЕНИЕ ДО-214. s: Упаковка / ящик: DO-214AA, SMB (3 вывода), Compak; Упаковка: лента и катушка (TR); Технология: смешанная технология; Приложения: SLIC; Количество цепей: 2; Напряжение — рабочее: 58В; Напряжение — зажим: 77 В; Мощность (Вт): -; Статус без содержания свинца: без содержания свинца; Статус RoHS :.
P1100SB : ТВ — тиристорная защита цепи 250А 130В; СИДАКТОР БИ 90В 250А ДО-214АА. s: Упаковка / футляр: DO-214AA, SMB; Упаковка: навалом; Емкость: 50 пФ; Напряжение — выключатель: 130В; Ток — пиковый импульс (8 x 20 с): 250 А; Ток — пиковый импульс (10 x 1000 с): 80 А; Статус без свинца: содержит свинец; Статус RoHS: не соответствует требованиям RoHS.
P3406UALRP : ТВ — тиристорная защита цепи 150А, 400В; SIDACTOR SYM 4CHP 300V 50A MS013. s: Упаковка / ящик: 6-SOIC (0.295 дюймов, ширина 7,50 мм); упаковка: лента и катушка (TR); емкость: 20 пФ, 30 пФ; напряжение — переключение: 400 В; ток — пиковый импульс (8 x 20 с): 150 А; ток — пиковый импульс (10 x 1000 с) : 45A; Бессвинцовый статус: Бессвинцовый; Статус RoHS: Соответствует RoHS.
3001 00020287 : Крепление на шасси для защиты цепи регулятора температуры; ТЕРМОСТАТ 3001 SER НЕГЕРМЕТИЧНЫЙ. s: Схема: -; Тип установки: Крепление на шасси; Температура переключения: -; Упаковка / ящик: цилиндр с монтажным фланцем; Статус без содержания свинца: без содержания свинца; Статус RoHS: Соответствует RoHS.
VP3225K401R275 : Варисторы 350 вольт 400A 3225. Варисторы KEMET SMD защищают электрические цепи от разрядов большой энергии. Каждая серия идеально подходит для целевого использования: s: Продукт: MLV; Тип завершения: SMD / SMT; Упаковка / ящик: 3225 (8063 метрическая система); Номинальное напряжение переменного тока: 275 В переменного тока; Номинальное напряжение постоянного тока: 350 В постоянного тока; Напряжение варистора: 430 В; Напряжение зажима: 710 В; Пиковый импульсный ток :.
Двухкомпонентная система PhoPR Clostridium acetobutylicum участвует в регуляции фосфат-зависимых генов
% PDF-1.4 % 46 0 объект > эндобдж 47 0 объект > поток ; изменено с помощью iText® 7.1.7 © 2000-2019 iText Group NV (версия AGPL) 2008-09-23T13: 25: 44Z2021-05-11T09: 10: 43-07: 002021-05-11T09: 10: 43-07 : 00XPPapplication / pdf
— CIAST
GCE) Уровень 1 (RS)Программа Pembelajaran Elektronik & Multimedia (PEM) (ICT)
29.03.2021 — 06.04.2021
INDUKSI SLDN KHAS ISEEDUCATION (SLDN_RS)
Программа Pembangunan Profesional Kemahiran (SPD) (Индивидуальная)
04.03.2021 — 04.03.2021
ПРОГРАММА ОБУЧЕНИЯ ТРЕНЕРОВ NDTS KHAS EADECO (COACH_SLDN)
Программа Pembangunan Profesional Kemahiran (SPD) (Индивидуальная)
04.05.2021 — 04.08.2021
ПРОГРАММА ОБУЧЕНИЯ ТРЕНЕРОВ NDTS WILAYAH UTARA (COACH_SLDN)
Программа Pembangunan Profesional Kemahiran (SPD) (Модульная)
04.05.2021 — 04.08.2021
ПРОГРАММА ОБУЧЕНИЯ ТРЕНЕРОВ NDTS WILAYAH SELATAN (TRAINER_SLDN)
Программа Pembangunan Profesional Kemahiran (SPD) (Модульная)
04.05.2021 — 04.09.2021
INDUKSI SLDN KHAS JABATAN PENJARA 01 (SLDN_RS)
Программа Pembangunan Profesional Kemahiran (SPD) (Индивидуальная)
05.04.2021 — 05.04.2021
ПРОГРАММА ОБУЧЕНИЯ ТРЕНЕРОВ NDTS KHAS PERODUA (COACH_SLDN)
Программа Pembangunan Profesional Kemahiran (SPD) (Индивидуальная)
04.05.2021 — 04.08.2021
TM001 / VTO: ПЕДАГОГИЧЕСКОЕ ОБУЧЕНИЕ / ПРОФЕССИОНАЛЬНОЕ ОБУЧЕНИЕ (МОДУЛЬНОЕ) -KBS
Программа Pembangunan Profesional Kemahiran (SPD) (Модульная)
04.05.2021 — 07.02.2021
INDUKSI SLDN KHAS KK SUPERMART (SLDN_RS)
Программа Pembangunan Profesional Kemahiran (SPD) (Индивидуальная)
04.06.2021 — 06.04.2021
INDUKSI SLDN KHAS JABATAN PENJARA 02 (SLDN_RS)
Программа Pembangunan Profesional Kemahiran (SPD) (Индивидуальная)
04.06.2021 — 06.04.2021
INDUKSI SLDN KHAS JABATAN PENJARA 03 (SLDN_RS)
Программа Pembangunan Profesional Kemahiran (SPD) (Индивидуальная)
04.07.2021 — 04.07.2021
ПРОГРАММА ОБУЧЕНИЯ ТРЕНЕРОВ NDTS CIAST1 (COACH_SLDN)
Программа Pembangunan Profesional Kemahiran (SPD) (Модульная)
04.07.2021 — 04.08.2021
Adobe Photoshop (модульный)
Программа Pembelajaran Elektronik & Multimedia (PEM) (ICT)
04.07.2021 — 04.08.2021
ТАКЛИМАТ БАХАН ПЕНГАДЖАРАН БЕРТУЛИС (ВИМ) ДАН СОАЛАН ЛАТИХАН КЕМАХИРАН СИРИ 1/2021
Программа Перанканган дан Пембангунан Латихан (PPL) (Пембангунан)
04.08.2021 — 04.08.2021
INDUKSI SLDN KHAS JABATAN PENJARA 04 (SLDN_RS)
Программа Pembangunan Profesional Kemahiran (SPD) (Индивидуальная)
04.08.2021 — 04.08.2021
INDUKSI SLDN KHAS HANDA INDAH (SLDN_RS)
Программа Pembangunan Profesional Kemahiran (SPD) (Индивидуальная)
09.04.2021 — 09.04.2021
INDUKSI SLDN KHAS JSS TAILORING 01 (SLDN_RS)
Программа Pembangunan Profesional Kemahiran (SPD) (Индивидуальная)
04.10.2021 — 10.04.2021
INDUKSI SLDN KHAS JSS TAILORING 02 (SLDN_RS)
Программа Pembangunan Profesional Kemahiran (SPD) (Индивидуальная)
10.04.2021 — 10.04.2021
INDUKSI SLDN KHAS JSS TAILORING 03 (SLDN_RS)
Программа Pembangunan Profesional Kemahiran (SPD) (Индивидуальная)
04.11.2021 — 04.11.2021
INDUKSI SLDN KHAS MAXIPURA (SLDN_RS)
Программа Pembangunan Profesional Kemahiran (SPD) (Индивидуальная)
11.04.2021 — 11.04.2021
ПРОГРАММА ОБУЧЕНИЯ ТРЕНЕРОВ NDTS ХАС ПРАСАРАНА (TRAINER_SLDN)
Программа Pembangunan Profesional Kemahiran (SPD) (Индивидуальная)
12.04.2021 — 15.04.2021
(PDF) जागतिकीकरणाचे धोरण व नेहरू विचारधारा
(Ежеквартальный, рецензируемый и индексируемый журнал с высоким импакт-фактором)
NAVJYOT / Vol.IX / Выпуск — II ISSN 2277-8063
КОЭФФИЦИЕНТ ВОЗДЕЙСТВИЯ — 6.013, IIJIF
www.navjyot.net
4
tkxfrdhdj.kkps /kksj.ko usg # fopkj / kkjk
frd000hd / tkjkfrd000hd usg # fopkj / kkjktkxfrdhdj.kkps /kksj.ko usg # fopkj / kkjktkxfrdhdj.kkps /kksj.ko usg # fopkj / kkjk
MkW- jfoanz 9000 Hkz. ] jkT; ‘kkL = vf / kfoHkkx] leUo; d] usg # vH; kl dsanz]
f’kokth fo | kihB] dksYgkiwj-
…………………………………………… ………………………………………….. ………………….
izLrkouk
izLrkoukizLrkouk
izLrkouk
iafMr tokgjyky usg # gs vlkekU;] izHkkoh yksdksÙkj Oks; ;
vkanksyukrhy usrk] Hkkjrkps iariz / kku], d eqRlÌh o vkarjjk ”Vªh; izfrek izfr ”Bk vl.kkjk
ekuorkoknh usrk] v’kk Hkwfedsrwu iafMr usg # aph vksG [k Hkkjrkykp uOgs] rj txkykgh
; ; kus nksu Hkkxkar foHkkxysyk
vkgs ifgY; k dkGkr Lokra «; izkIrhlkBh> xM.kkjs usg #] rj nqljk dky [kaM Eg.kts
Lokra «; ksÙkj dkGkr Hkkjrkyk txkP; k ikBhoj, d uos yksd’kkgh jk» Vª? kMfo.; #ikpk gksrk- iafMr usg # auk tso R; kauk fojks / kgh lgu djkok ykxyk- usg # aP; k /kksj.kkps leFkZd vkf3. из [kji.ks usg # fopkjkaph fpdhRlk djrp vkgsr- HkkjrkP; k jktdkj.kkr] lektdkj.kkr usg # aps vrqyuh; ; ksxnku vkgs- vkiY; k thoukrhy rCcy rhl o ”ksZ iafMr usg # auh Hkkjrh; Локра «; izkIrhlkBh fczfV’kka’kh> xM.; ? kkyfoys- Hkkjrh; Локра «; vkanksyukr fczfV’k lÙks’kh> xMr vlrkuk, danjhr; k dky [kaMkr iafMr usg # auk toGikl uÅ o ”kkZpk dkjkokl Hkksxyk- Hkkjrh; Lokra «; izkIrhuarj iafMr tokgjyky usg # auh Hkkjrkps usr`Ùo lrjk o” ksZ dsys-; k dkGkr Hkkjrkph vk / kqfud n`’k; ; egÙoiw.kZ vkf.k vrqyuh; vlsp vkgs- ns’kkrhy loZlkekU; tursr vkRefo’okl fuekZ.k d # u rks ok usr`Ùokpkp, d Hkkx gksrk- Hkkjrkph vksG [k [kÚ; k vFkkZus u nskyk d #.kkjs gs, d egku usrs gksrs- Hkkjrkyk vkf.k tkxfrd jktdkj.kkyk usg # fopkjkapk, d izokg vtwugh izHkkfor o ekxZn’gkzjk fükkzykkjkjkjkjkjkjkjkjkjkjkfjkjkjkjkjkjkjkjkjkjkjkjkjkjkjkjkjkjkjkjkjkjkf? rj usg # fojks / kh fopkjkaph nqljh cktwlq / nk vki.kkl VkGwu pky.kkj ukgh- R; keqGs iafMr tokgjyky usg # aps usr`Ùo oknjfk. oknkeqGs usg # AP; k usr`Ùokps; ksxnku vkiY; kyk VkGrk; sr ukgh- R; keqGsp usg # AP; k usr`Ùokph] R; kaP; k ; ksxnk ppkf rVLF.k iafMr tokgjyky usg # AP; k cíy vlysY; k let & xSjlet; akps fujkdj.k vktP; k ifjfLFkrhr gks.ks xjtsps vkgs- iafMr tokg`jykHykskr. kwu usg # iz.khr lalnh; yksd’kkghph mHkkj.kh Hkkjrkr lqn` [kk.kkjs rÙo Hkk’jrks kf; kkr #tw ‘kdys] brdsp uOgs] rj rs fodflr gksÅu Hkkjrkr le` / n gksÅ’ kdys vkf.k R; koj Hkkjrh; ‘KkluO; oLFkk fLFkj gksowu fLFkjkow‘ kdyh- usg # AP; k Hkkjrkrhy usr`Ùokps vkf.k dk; kZps ewY; ekiu djko; kps EgVys rj Hkkjrkr dsysY; k dk; kZps ikjMs fuf’prp tM Bjysys vkgs- gs vki.kkl fufoZokniL.ks-koskU; турсык вкиЫ; клексжы вкОгкукаф лксМо.квд гох гксрх вкф.к рс йодж лксМфо.кс ‘кД; uOgrs- R; kl Bjkohd dky [каМ tkÅ ns.ks xjtsps gksrs- usg # auk [kk = h gksrh dh; kstukc / nfjR; k iz; Ru d # u gs iz’u lksMfork ; srkr] klkBh txkr nh? kZdkG ‘kkarark ukan.ks xjtsps gksrs (I.k rh ‘kkarrk Hkkjrkyk ykHkyh ukgh-; k dkGkr usg # auh fodklØe vk [kr vk / kqfud Hkkjrkph ik; kojp fVdkdkj foospu djrkr- rls u djrk usg # AP; k fopkjkaph ppkZ] ewY; ekiu; Fkklkaxi.ks Ogkos]; kdfjrk uO; kus fopkjeaFku Фосфорсодержащие биомолекулы участвуют в широком спектре клеточной активности, включая обработку информации, энергетический метаболизм, передачу сигналов, регуляцию активности белков и поддержание кислотно-основного гомеостаза.Кроме того, клеточные оболочки некоторых бактерий (например, Bacilli и Staphylococci ) содержат липотейхоевую кислоту (LTA) и стеночную тейхоевую кислоту (WTA), анионные полимеры с высоким содержанием фосфора. LTA представляет собой полимер глицеринфосфата, который проникает в клеточную стенку из липида, закрепленного в клеточной мембране. WTA представляет собой полимер глицерин- или рибитолфосфата, ковалентно связанный с пептидогликаном (Weidenmaier and Peschel, 2008; Percy and Gründling, 2014). Эти анионные полимеры играют важную роль в клеточной морфологии и делении клеток (Swoboda et al., 2010; Перси и Грюндлинг, 2014). Следовательно, достаточное количество фосфора является предпосылкой для жизнеспособности клеток. Это проблема для бактерий, особенно тех, которые обитают в местах обитания, таких как почва, где уровень свободного фосфата может быть низким из-за pH-зависимого образования осадков (Batjes, 1997). Бактерии применяют две общие стратегии для поддержания адекватного поступления фосфора: (i) внутриклеточное накопление и (ii) удаление фосфата с помощью ферментов, индуцируемых в условиях ограничения фосфора.Многие бактерии хранят фосфор в виде полифосфата, полимера из сотен фосфатных остатков, связанных высокоэнергетическими фосфоангидридными связями, который синтезируется полифосфаткиназой (PPK1) с использованием нуклеотидтрифосфатов (Brown and Kornberg, 2008; Rao et al., 2009; Jiménez et al. , 2017). Фосфат высвобождается из хранилища полифосфатов в результате разложения экзо- и эндо-полифосфатфосфатазами (Brown and Kornberg, 2008; Achbergerová and Nahálka, 2011). Однако Bacillus subtilis подвид subtilis (далее именуемый B.subtilis ) не может синтезировать полифосфат, но вместо этого использует WTA в качестве хранилища, из которого фосфат высвобождается за счет комбинированной активности фосфодиэстераз GlpQ и PhoD, экспрессия которых увеличивается при ограничении фосфата в зависимости от PhoPR (Eder et al., 1996; Antelmann et al., 2000; Myers et al., 2016). Адаптация к ограничению фосфатов часто опосредуется двухкомпонентной системой передачи сигналов (TCS) под названием PhoBR в Escherichia coli и PhoPR в B.subtilis (далее называемый ответом PHO). PhoR представляет собой сенсорную киназу, активность которой зависит от доступности фосфата. При активации PhoR фосфорилирует свой родственный регулятор ответа (PhoB или PhoP), который, в свою очередь, управляет программой экспрессии генов, чтобы адаптировать бактерии к состоянию ограниченного фосфата. PHO-ответы этих бактерий характеризуются (i) повышенной экспрессией и секрецией ферментов, которые поглощают фосфат; (ii) повышенная экспрессия высокоаффинного транспортера фосфата ABC-типа (PstSCAB) и (iii) усиление ответа за счет положительной ауторегуляции оперонов phoBR и phoPR . Как PhoR воспринимает фосфат и как его активность регулируется в зависимости от доступности фосфата, было исследовано в основном на E. coli и B. subtilis . Появились два различных механизма, которые контролируют баланс между активностями автокиназы и фосфатазы PhoR. Первый механизм, проиллюстрированный активацией PhoBR в E. coli , реагирует на транспорт фосфата и опосредуется транспортером PstSCAB и адаптерным белком PhoU (Hsieh and Wanner, 2010).Второй механизм, представленный PhoPR в B. subtilis, , отвечает на метаболизм WTA и опосредуется промежуточными соединениями в биосинтезе WTA (Botella et al., 2014; Prunty et al., 2018). Наблюдается изменение обоих механизмов, которые позволяют бактериям обеспечивать достаточное количество фосфора в своих конкретных экологических нишах. Семь белков необходимы и достаточны для индукции ответа PHO у E.coli : PhoBR TCS, транспортер фосфата PstSCAB ABC-типа и белок PhoU, который кодируется в опероне pstSCABphoU (Hsieh and Wanner, 2010). PhoBR — это классический TCS. PhoR связан с мембраной и состоит из двух трансмембранных доменов и цитоплазматического домена PAS (Per Arndt Sim), который взаимодействует с PhoU. Регулятор ответа PhoB активируется посредством PhoR-опосредованного фосфорилирования и связывается с PHO-боксами, расположенными в промоторной области генов регулона PhoBR (Blanco et al., 2002). Транспортер PstSCAB ABC-типа состоит из периплазматически расположенного белка PstS, который связывает фосфат, белков PstCA, расположенных внутри мембраны, которые образуют транспортный канал, и димера PstB, который связывает АТФ и обеспечивает энергией транспортный процесс. Белок PhoU представляет собой связанный с мембраной металл-связывающий белок, состоящий из двух трех-альфа-спиральных пучков, которые образуют мультимеры (Liu et al., 2005; Oganesyan et al., 2005). Генетический анализ показал, что ответ PHO конститутивно активируется, когда либо транспортер PstSCAB, либо PhoU удаляется (Figure 1A; Hsieh and Wanner, 2010).Это означает, что PhoR находится в режиме автокиназы по умолчанию в условиях ограничения фосфата и что транспортер с высоким сродством PstSCAB и PhoU оба необходимы для его преобразования в режим фосфатазы в условиях избытка фосфата (Hsieh and Wanner, 2010). Белок PhoU взаимодействует с доменом PAS PhoR и белком PstB транспортера PstSCAB с образованием связанного с мембраной сигнального комплекса, который преобразует PhoR в режим активности фосфатазы (Gardner et al., 2014, 2015). Белок PhoU также модулирует активность транспортера PstSCAB (Rice et al., 2009). Результаты показывают, что баланс между автокиназным и фосфатазным режимами активности PhoR не контролируется ни концентрацией внутриклеточного фосфата, ни транспортом фосфата per se (Cox et al., 1988, 1989; Rao et al., 1993). Вместо этого есть данные, полученные с помощью генетического подхода, стабилизирующего PstB в двух различных конформациях, что конформационные изменения в транспортере PstSCAB важны для определения баланса между автокиназным и фосфатазным режимами активности PhoR (Vuppada et al., 2018). Когда транспортер присутствует в обращенной наружу «закрытой» структуре, сигнальный комплекс PstSCAB / PhoU либо не взаимодействует с PhoR (рис. 1A, сплошные линии PhoU), либо взаимодействует с PhoR (рис. 1A, пунктирные линии PhoU), способствуя активности автокиназы. , состояние, которое существует в условиях ограничения фосфата. Однако, когда переносчик присутствует в обращенной внутрь «открытой» структуре, сигнальный комплекс PstSCAB / PhoU взаимодействует с PhoR, чтобы способствовать активности фосфатазы, состоянию, которое существует в условиях достаточности фосфата (рис. 1B).Таким образом, наличие фосфата в E. coli определяется косвенно через конформационные изменения в транспортере PstSCAB (Gardner et al., 2015; Vuppada et al., 2018). Фактически, теперь было показано, что переносчики играют сигнальную роль в контроле активности нескольких TCS (обзоры см. Tetsch and Jung, 2009; Piepenbreier et al., 2017). РИСУНОК 1. Модель для контроля баланса между активностями автокиназы PhoR и фосфатазы в E.coli , генерируемые конформационными изменениями в транспортере фосфата PstSCAB. Белок PhoR может существовать либо в режиме автокиназы в условиях ограничения фосфата (A) , либо в режиме фосфатазы в условиях достаточности фосфата (B) . Предполагается, что эти альтернативные состояния определяются различными конформациями транспортера PstSCAB, которые передаются PhoR с помощью PhoU для определения его активности. Когда PstSCAB находится в закрытой обращенной наружу конформации (A) , адаптерный белок PhoU либо не может взаимодействовать с PhoR (сплошные линии PhoU), либо взаимодействует с доменом PAS PhoR, способствуя активности автокиназы (пунктирные линии PhoU) в определенных условиях. ограничения фосфата.Родственный регулятор ответа PhoB фосфорилируется в этом состоянии. Однако, когда PstSCAB находится в открытой обращенной внутрь конформации (B) , адаптерный белок PhoU взаимодействует с доменом PAS PhoR, чтобы способствовать активности фосфатазы в условиях достаточности фосфата. Родственный регулятор ответа PhoB дефосфорилируется в этом состоянии. Эта диаграмма представляет собой адаптацию моделей и рисунков, представленных в работах Hsieh and Wanner (2010) и Vuppada et al. (2018). Тема регуляции PhoBR / PstSCAB / PhoU широко используется для контроля метаболизма фосфатов и связанных с ним клеточных процессов (например,g., вирулентность) у бактерий (Lamarche et al., 2008; Chekabab et al., 2014; Santos-Beneit, 2015). Тем не менее, тема может быть изменена путем изменения количества, активности или взаимодействия составляющих компонентов, как показано на этих выбранных примерах. В Caulobacter crescentus PhoU является важным белком, который не контролирует активность PhoR, но регулирует внутриклеточный метаболизм фосфата (Lubin et al., 2015). Есть два белка PhoU, кодируемых в Staphylococcus epidermidis : ген phoU1 расположен в опероне pstSCABphoU1 , а phoU2 расположен перед геном, гомологичным переносчику фосфата PitA (Wang et al., 2017). Генетический анализ показывает, что только PhoU2 контролирует активность PhoPR (Wang et al., 2017). Streptococcus pneumoniae кодирует единую двухкомпонентную систему PnpRS, расположенную перед транспортером фосфата Pst1 ( pstS1C1A1B1phoU1 ), но кодирует второй транспортер фосфата Pst2 ( pstS2C2A2B2phoU2) в отдельном месте. Экспрессия Pst2 является конститутивной, а PhoU2 ингибирует активность транспортера Pst2. Экспрессия Pst1 индуцируется ограничением фосфата PnpRS-зависимым образом, тогда как PhoU1 ингибирует активность транспортера Pst1 (Zheng et al., 2016). Однако PhoU2, но не PhoU1, контролирует активность PnpRS аналогично тому, как это происходит в E. coli (Zheng et al., 2016). Эти отобранные примеры показывают универсальность регуляторной темы PhoBR / PstSCAB / PhoU и показывают, как она может варьироваться у бактерий, чтобы реагировать на доступность фосфата в их конкретной экологической нише. Некоторые особенности ответа PHO в B.subtilis указывают на то, что он отличается от такового у E. coli . Геном B. subtilis не кодирует гомолог PhoU, и реакция PHO обычно индуцируется в штамме с удаленным высокоаффинным транспортером фосфата ( pstSCAB 1 B 2 ) (Qi et al., 1997 ). Кроме того, в отличие от E. coli , состав и метаболизм анионных полимеров клеточной стенки изменяются PhoPR-зависимым образом во время реакции PHO в B.subtilis (Liu, Hulett, 1998; Liu et al., 1998; Allenby et al., 2005; Botella et al., 2011, 2014; Salzberg et al., 2015). При активации PhoP∼P репрессирует транскрипцию оперона tagAB , тем самым уменьшая синтез WTA, и активирует транскрипцию оперона tuaA-H , тем самым увеличивая синтез тейхуроновой кислоты, замещающего нефосфатсодержащего анионного полимера (Рисунок 2; Лю и Hulett, 1998; Liu et al., 1998; Allenby et al., 2005; Botella et al., 2011).Кроме того, гены регулона PhoPR кодируют множество ферментов, поглощающих фосфат, включая посфодиэстеразы GlpQ и PhoD, которые участвуют в деградации WTA (Eder et al., 1996; Antelmann et al., 2000; Allenby et al., 2005; Botella et al. ., 2011; Myers et al., 2016). Кроме того, B. subtilis не может синтезировать полифосфат. РИСУНОК 2. Модель для контроля баланса между активностями автокиназы PhoR и фосфатазы в B.subtilis и B. spizizenii , генерируемые промежуточными продуктами метаболизма тейхоевой кислоты в стенке. В B. subtilis и B. spizizenii предполагается, что автокиназный и фосфатазный режимы активности PhoR определяются уровнями биосинтетических промежуточных продуктов WTA. (A) Метаболический путь биосинтеза WTA в B. spizizenii показан с конечным продуктом липида V, состоящим из поли (рибитолфосфата), присоединенного к липидному носителю (зубчатые линии) линкером, состоящим из модифицированного пирофосфата. сахара (Glc N Ac, Man N Ac) и одиночный глицеринфосфатный фрагмент.Геномная организация биосинтетических генов показана в рамке. Предполагается, что ферментный продукт TarA или TarB активирует активность автокиназы PhoR (стрелки с пунктирными линиями указывают на предполагаемое взаимодействие). Уровень активирующего промежуточного соединения будет определяться балансом между синтезом WTA и тейхуроновой кислоты в ограниченных фосфатом клетках из-за взаимной взаимосвязи между этими двумя метаболическими путями. (B) Метаболический путь биосинтеза WTA в B.subtilis показан с конечным продуктом липида V, состоящим из поли (глицеринфосфата). Геномная организация биосинтетических генов (в рамке) показывает, что оперон tagAB репрессируется активированным PhoP∼P (-). Предполагается, что ферментный продукт TagA или TagB активирует активность автокиназы PhoR (стрелки с пунктирными линиями). Однако в B. subtilis липид V, состоящий из поли (глицеринфосфата), может ингибировать активность автокиназы PhoR (показано in vitro, синяя линия с блоком).Таким образом, баланс между активностями автокиназы PhoR и фосфатаз будет определяться уровнем активирующих промежуточных продуктов (TagA или TagB, черные стрелки), степенью активации синтеза тейхуроновой кислоты и подавления синтеза WTA с помощью PhoP∼P и уровень ингибирующего липида V, состоящего из поли (глицеринфосфата), синяя линия с блоком). PhoR в B. subtilis также отличается от такового в E. coli наличием внецитоплазматической петли, способной образовывать домен PAS, который не требуется для индукции ответа PHO в лабораторных условиях (Рисунок 2; Shi and Hulett, 1999; Чанг и др., 2010; Ботелла и др., 2014). Усиление ответа PHO за счет позитивной ауторегуляции транскрипции phoPR усиливается связыванием PhoP∼P с PhoP-боксами, расположенными между стоп-кодоном phoR и терминатором оперона, по еще неизвестному механизму (Salzberg et al., 2015 ). Поэтому, возможно, неудивительно, что активация реакции PHO у B. subtilis отличается от таковой у E. coli и что она включает метаболизм WTA. Связь между метаболизмом анионных полимеров и активностью PhoR была установлена в г.subtilis , обнаружив, что амплификация ответа PHO задерживается и ослабляется в клетках, неспособных синтезировать тейхуроновую кислоту (Botella et al., 2014). Поскольку транскрипция оперона tuaA-H требует активированного PhoP∼P, этот результат указывает на то, что биосинтез тейхуроновой кислоты усиливает ответ PHO с помощью механизма положительной обратной связи, чтобы увеличить активность автокиназы PhoR (Botella et al., 2014). Это наблюдение особенно важно, поскольку синтез WTA снижается за счет опосредованной PhoP∼P репрессии экспрессии tagAB .Кроме того, существует обратная связь между синтезом WTA и тейхуроновой кислоты в B. subtilis (продукт фермента TagO является предшественником в обоих метаболических путях), подразумевая, что увеличение синтеза тейхуроновой кислоты вызовет снижение синтеза WTA (рис. ; Botella et al., 2014). Таким образом, мы стремились установить, вызвана ли повышенная активность PhoR во время амплификации реакции PHO повышением уровня промежуточного продукта биосинтеза тейхуроновой кислоты, который способствует активности аутокиназы, или снижением уровня промежуточного продукта биосинтеза WTA, который ингибирует активность аутокиназы. .Генетические и биохимические данные показывают, что (i) ослабление реакции PHO у штаммов, неспособных синтезировать тейхуроновую кислоту, может быть подавлено сопутствующим снижением синтеза WTA и (ii) активность автокиназы PhoR ингибируется in vitro производным WTA промежуточный продукт биосинтеза, продуцируемый ферментом TagF (Botella et al., 2014). Эти данные показывают, что активность автокиназы PhoR реагирует на метаболизм WTA в B. subtilis , который ингибируется продуктом фермента TagF (Botella et al., 2014). Поскольку ответ PHO усиливается у B. subtilis с помощью двух процессов, требующих активации PhoP∼P (т.е. снижения синтеза WTA и активации синтеза тейхурона), очевидно, что он должен быть инициирован другим PhoP∼P. -независимый механизм. Таким образом, инициация PHO-ответа была направлена на Bacillus subtilis подвид spizizenii (далее именуемый B. spizizenii ), который синтезирует WTA, состоящий из поли (рибитолфосфата) ( tarABIJKL генов) (Prunty et al., 2018). Изучение ответа PHO у B. spizizenii имеет то преимущество, что он не амплифицируется, как в B. subtilis [т.е. промежуточные соединения WTA, состоящие из поли (рибитолфосфата), не ингибируют активность автокиназы PhoR], и экспрессия WTA биосинтетические ферменты не репрессируются PhoP∼P (Prunty et al., 2018). Три доказательства указывают на то, что ответ PHO активируется промежуточным звеном в синтезе WTA у B. spizizenii . (1) В штамме с индуцибельной экспрессией оперона tarABIJKL , кодирующего ферменты биосинтеза WTA, величина реакции PHO прямо пропорциональна уровню добавленного индуктора (Prunty et al., 2018). (2) PHO-ответ повышен у штамма B. spizizenii , неспособного синтезировать тейхуроновую кислоту, что указывает на то, что начало синтеза тейхуроновой кислоты снижает уровень интермедиата WTA, который активирует PHO-ответ (Prunty et al., 2018 ). (3) Повышенный ответ PHO у штамма, неспособного синтезировать тейхуроновую кислоту, снижается за счет снижения экспрессии генов, кодирующих ферменты биосинтеза WTA (Prunty et al., 2018). Аналогичные результаты были получены в B.spizizenii , экспрессирующие либо гомологичную киназу PhoR, либо киназу PhoR из B. subtilis , что свидетельствует о том, что активация происходит по одному и тому же механизму у обоих подвидов (Prunty et al., 2018). Эти результаты предполагают, что промежуточное соединение в синтезе WTA активирует активность аутокиназы PhoR как у B. subtilis , так и у B. spizizenii (Prunty et al., 2018). Поскольку только ферментативные стадии TagA / TarA и TagB / TarB биосинтетических путей WTA являются общими для B.subtilis и B. spizizenii , мы пришли к выводу, что тот или иной их ферментный продукт способствует активности аутокиназы PhoR (Prunty et al., 2018). Модель активации реакции PHO у B. spizizenii и B. subtilis (рисунки 2A, B, соответственно) предполагает, что при ограничении фосфата (i) наблюдается всплеск клеточного уровня промежуточных продуктов WTA, вызванный снижение скорости роста и снижение синтеза клеточной стенки и (ii) повышенный уровень ферментного продукта TagA / TarA или TagB / TarB способствует активности автокиназы PhoR (Prunty et al., 2018). Кроме того, активность автокиназы PhoR контролируется в B. subtilis только уровнем ферментного продукта TagF, состоящего из поли (глицеринфосфата) (рис. 2B; Botella et al., 2014). Таким образом, баланс активности PhoR аутокиназы и фосфатазы зависит от метаболизма WTA в B. subtilis и B. spizizenii (Prunty et al., 2018). Кинетика PHO-ответов B.subtilis и B. spizizenii различаются (Botella et al., 2014; Prunty et al., 2018). У B. subtilis реакция PHO активируется, затем усиливается и поддерживается, пока сохраняется ограничение по фосфату (Botella et al., 2014). У B. spizizenii реакция PHO активируется, но затем постепенно отключается, даже при сохранении условий ограничения фосфата (Prunty et al., 2018). Эти отдельные ответы не связаны с различиями в киназах PhoR, которые являются высокогомологичными и экспериментально доказанными функционально эквивалентными (Prunty et al., 2018). Вместо этого различные ответы PHO происходят из того факта, что продукт фермента TagF, состоящий из поли (глицеринфосфата), ингибирует активность автокиназы PhoR в B. subtilis , но соответствующий продукт фермента TarL, состоящий из поли (рибитолфосфата), не ингибирует автокиназу PhoR активность в B. spizizenii (рисунок 2; Botella et al., 2014; Prunty et al., 2018). Следовательно, особенности хозяина, которые влияют на экспрессию ферментов биосинтеза WTA и уровень промежуточных соединений WTA, будут влиять на ответы PHO этих двух подвидов.Эти особенности хозяина включают геномную организацию биосинтетических генов WTA, регуляцию их экспрессии и их клеточного уровня мРНК (Prunty et al., 2018). Все ферменты биосинтеза WTA кодируются одним опероном ( tarABIJKL ) в B. spizizenii (рис. 2A), но разделены на два оперона ( tagAB и tagDEF ) в B. subtilis (рис. 2B). . Экспрессия оперона tarABIJKL не регулируется PhoPR в B.spizizenii (фиг. 2A), тогда как экспрессия оперона tagAB негативно регулируется активированным PhoP∼P в B. subtilis (фиг. 2B). В B. spizizenii уровни мРНК генов tarABIJKL равномерно снижены до ~ 15% от уровня, обнаруженного в экспоненциально растущих клетках с высоким содержанием фосфатов (Prunty et al., 2018). Однако в B. subtilis уровни мРНК генов tagAB снижены до <1%, в то время как уровень tagF снижен только до 27% от уровня, обнаруженного в экспоненциально растущих, переполненных фосфатом клетках.Таким образом, эти разные хозяева имеют прямые различные ответы PHO в B. subtilis и B. spizizenii путем изменения метаболизма WTA, что позволяет оптимизировать адаптацию к доступности фосфата в определенных экологических сообществах. Бактериальный ответ PHO широко сохраняется среди бактерий. Однако появились два разных механизма, с помощью которых активность PhoR контролируется способом, который зависит от доступности фосфата.Механизм, реагирующий на конформацию транспортера фосфата PstSCAB в сочетании с PhoU, примером которого является PhoBR в E. coli , широко распространен среди грамположительных и грамотрицательных бактерий. Отдельный механизм, который реагирует на синтез WTA, представлен на примере PhoPR в B. subtilis . Непонятно, почему у B. subtilis развился такой отчетливый механизм, но он может относиться к высокофосфатному составу анионных полимеров клеточной стенки, способности изменять анионный полимерный состав во время ограничения фосфата, неспособности синтезировать полифосфат и использование WTA в качестве хранилища фосфатов.Вероятно, что PhoPR активируется в механизме типа PhoU у стафилококков, что позволяет предположить, что фосфорсодержащий WTA не является определяющим признаком в эволюции нового механизма у B. subtilis . Однако различия в доступности фосфора в их естественных привычках могли сыграть значительную роль, а его доступность в почве была особенно проблематичной для видов Bacillus. Поэтому будет интересно установить, как окраски B. subtilis и Staphylococcus aureus , экспрессирующие механизм активации PhoPR другой бактерии, адаптируются к ограничивающим фосфат условиям и выживают в своей естественной среде обитания. К.Д. оформил рецензию и подготовил рукопись. Эти проекты финансировались Ирландским научным фондом, награды главного исследователя 08 / IN.1 / B1859 и 12 / 1A / 1570, выданные KD. Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов. Автор благодарит К.М., Майкла Пранти, Джоан Геогеган и Эбигейл О’Брайен за критическое прочтение рукописи. Алленби, Н. Э., О’Коннор, Н., Прагай, З., Уорд, А. К., Випат, А., и Харвуд, К. Р. (2005). Полногеномный транскрипционный анализ стимула фосфатного голодания Bacillus subtilis. J. Bacteriol. 187, 8063–8080. DOI: 10.1128 / JB.187.23.8063-8080.2005 PubMed Аннотация | CrossRef Полный текст | Google Scholar Антельманн, Х., Шарф, К., Хеккер, М. (2000). Белки, индуцируемые фосфатным голоданием Bacillus subtilis : протеомика и транскрипционный анализ. J. Bacteriol. 182, 4478–4490. DOI: 10.1128 / JB.182.16.4478-4490.2000 PubMed Аннотация | CrossRef Полный текст | Google Scholar Батьес, Н. Х. (1997). Мировой набор данных по производным свойствам почвы, подготовленный почвенным отделом ФАО-ЮНЕСКО для глобального моделирования. Управление использованием почвы. 13, 9–16. Google Scholar Бланко, А. Г., Сола, М., Гомис-Рют, Ф. Х. и Колл, М. (2002). Распознавание тандемной ДНК с помощью PhoB, двухкомпонентного активатора транскрипции сигнальной трансдукции. Структура 10, 701–713. DOI: 10.1016 / S0969-2126 (02) 0 0761-X PubMed Аннотация | CrossRef Полный текст | Google Scholar Ботелла, Э., Дивайн, С. К., Хубнер, С., Зальцберг, Л. И., Гейл, Р. Т., Браун, Э. Д. и др. (2014). Активность автокиназы PhoR контролируется промежуточным звеном в метаболизме тейхоевой кислоты в стенке, которое ощущается внутриклеточным доменом PAS во время реакции ограничения фосфата, опосредованной PhoPR, у Bacillus subtilis . Мол. Microbiol. 94, 1242–1259. DOI: 10.1111 / mmi.12833 PubMed Аннотация | CrossRef Полный текст | Google Scholar Botella, E., Hübner, S., Hokamp, K., Hansen, A., Bisicchia, P., Noone, D., et al. (2011). Экспрессия гена клеточной оболочки в ограниченных фосфатом клетках Bacillus subtilis . Микробиология 157, 2470–2484. DOI: 10.1099 / mic.0.049205-0 PubMed Аннотация | CrossRef Полный текст | Google Scholar Чанг, К., Тесар, К., Гу, М., Бабнигг, Г., Иоахимиак А., Поккулури П. Р. и др. (2010). Внецитоплазматические PAS-подобные домены обычны в белках сигнальной трансдукции. J. Bacteriol. 192, 1156–1159. DOI: 10.1128 / JB.01508-09 PubMed Аннотация | CrossRef Полный текст | Google Scholar Чекабаб, С. М., Джубелин, Г., Дозуа, К. М., и Харел, Дж. (2014). PhoB активирует факторы вирулентности Escherichia coli O157: H7 в ответ на ограничение неорганических фосфатов. PLoS One 9: e94285. DOI: 10.1371 / journal.pone.0094285 PubMed Аннотация | CrossRef Полный текст | Google Scholar Кокс, Г. Б., Уэбб, Д., Годовац-Циммерманн, Дж., И Розенберг, Х. (1988). Arg-220 белка PstA необходим для транспорта фосфата через фосфат-специфическую транспортную систему в Escherichia coli , но не для репрессии щелочной фосфатазы. J. Bacteriol. 170, 2283–2286. DOI: 10.1128 / jb.170.5.2283-2286.1988 PubMed Аннотация | CrossRef Полный текст | Google Scholar Кокс, Г.Б., Уэбб Д. и Розенберг Х. (1989). Определенные аминокислотные остатки в белках PstB и PstC необходимы для транспорта фосфата системой Escherichia coli Pst. J. Bacteriol. 17, 1531–1534. DOI: 10.1128 / jb.171.3.1531-1534.1989 PubMed Аннотация | CrossRef Полный текст | Google Scholar Эдер С., Ши Л., Дженсен К., Ямане К. и Хьюлетт Ф. М. (1996). Bacillus subtilis секретируемая фосфодиэстераза / щелочная фосфатаза является продуктом гена регулона Pho, phoD. Микробиология 142, 2041–2047. DOI: 10.1099 / 13500872-142-8-2041 PubMed Аннотация | CrossRef Полный текст | Google Scholar Гарднер, С. Г., Джонс, К. Д., Таннер, Р., и МакКлири, В. Р. (2014). Белок PhoU из Escherichia coli взаимодействует с PhoR, PstB и металлами с образованием фосфатно-сигнального комплекса на мембране. J. Bacteriol. 196, 1741–1752. DOI: 10.1128 / JB.00029-14 PubMed Аннотация | CrossRef Полный текст | Google Scholar Гарднер, С.Г., Миллер, Дж. Б., Дин, Т., Робинсон, Т., Эриксон, М., Ридж, П. Г. и др. (2015). Генетический анализ, структурное моделирование и анализ прямого связывания подтверждают механизм передачи сигналов фосфата в Escherichia coli . BMC Genet. 16 (Приложение 2): S2. DOI: 10.1186 / 1471-2156-16- S2-S2 PubMed Аннотация | CrossRef Полный текст | Google Scholar Ламарш, М.Г., Ваннер, Б.Л., Крепен, С., и Харел, Дж. (2008). Фосфатный регулон и бактериальная вирулентность: регуляторная сеть, связывающая фосфатный гомеостаз и патогенез. FEMS Microbiol. Rev. 32, 461–473. DOI: 10.1111 / j.1574-6976.2008.00101.x PubMed Аннотация | CrossRef Полный текст | Google Scholar Лю В. и Хьюлетт Ф. М. (1998). Сравнение связывания PhoP с промотором tuaA со связыванием PhoP с другими промоторами Pho-регулона позволяет установить сайт связывания ядра Bacillus subtilis Pho. Микробиология 144, 1443–1450. DOI: 10.1099 / 00221287-144-5-1443 PubMed Аннотация | CrossRef Полный текст | Google Scholar Лю В., Эдер, С., Хьюлетт, Ф. М. (1998). Анализ экспрессии tagAB Bacillus subtilis и tagDEF во время фосфатного голодания определяет репрессорную роль для PhoP-P. J. Bacteriol. 180, 753–738. PubMed Аннотация | Google Scholar Лю Дж., Лу Ю., Йокота Х., Адамс П. Д., Ким Р. и Ким С. Х. (2005). Кристаллическая структура гомолога белка PhoU: новый класс металлопротеинов, содержащий многоядерные кластеры железа. J. Biol. Chem. 280, 15960–15966.DOI: 10.1074 / jbc.M414117200 PubMed Аннотация | CrossRef Полный текст | Google Scholar Любин, Э.А., Генри, Дж. Т., Фибигм, А., Кроссон, С., и Лауб, М. Т. (2015). Идентификация регулона PhoB и роли PhoU в реакции на фосфатное голодание Caulobacter crescentus . J. Bacteriol. 198, 187–200. DOI: 10.1128 / JB.00658-15 PubMed Аннотация | CrossRef Полный текст | Google Scholar Майерс, К. Л., Ли, Ф. К., Ку, Б.М., Эль-Халфави, О. М., Френч, С., Гросс, К. А. и др. (2016). Идентификация двух гидролаз стенки тейхоевой кислоты, индуцированной фосфатным голоданием, дает первое представление о пути деградации ключевого компонента бактериальной клеточной стенки. J. Biol. Chem. 291, 26066–26082. DOI: 10.1074 / jbc.M116.760447 PubMed Аннотация | CrossRef Полный текст | Google Scholar Оганесян В., Оганесян Н., Адамс П. Д., Джанчарик Дж., Йокота Х. А., Ким Р. и др. (2005). Кристаллическая структура «PhoU-подобного» регулятора поглощения фосфата из Aquifex aeolicus . J. Bacteriol. 187, 4238–4244. DOI: 10.1128 / JB.187.12.4238-4244.2005 PubMed Аннотация | CrossRef Полный текст | Google Scholar Перси М. Г., Грюндлинг А. (2014). Синтез и функция липотейхоевой кислоты у грамположительных бактерий. Annu. Rev. Microbiol. 68, 81–100. DOI: 10.1146 / annurev-micro-091213-112949 PubMed Аннотация | CrossRef Полный текст | Google Scholar Прунти, М. П., Ноун, Д., Дивайн, К. М. (2018). Разные PhoPR-опосредованные ответы на ограничение фосфата у Bacillus subtilis подвидов subtilis и spizizenii происходят из-за различий в составе и метаболизме тейхоевой кислоты в стенке. Мол. Microbiol. 109, 23–40. DOI: 10,1111 / мми. 13965 PubMed Аннотация | CrossRef Полный текст | Google Scholar Ци Ю., Кобаяши Ю. и Хьюлетт Ф. М. (1997). Оперон pst Bacillus subtilis имеет промотор, регулируемый фосфатом, и участвует в транспорте фосфата, но не в регуляции фосрегулона. J. Bacteriol. 179, 2534–2539. DOI: 10.1128 / jb.179.8.2534-2539.1997 PubMed Аннотация | CrossRef Полный текст | Google Scholar Рао, Н.Н., Гомес-Гарсия, М. Р. и Корнберг А. (2009). Неорганический полифосфат: необходим для роста и выживания. Annu. Rev. Biochem. 78, 605–647. DOI: 10.1146 / annurev.biochem.77.083007.093039 CrossRef Полный текст | Google Scholar Рао Н. Н., Робертс М. Ф., Торриани А. и Яшфе Дж. (1993). Влияние мутаций glpT и glpD на экспрессию гена phoA в Escherichia coli . J. Bacteriol. 175, 74–79. DOI: 10.1128 / jb.175.1.74-79.1993 PubMed Аннотация | CrossRef Полный текст | Google Scholar Райс, К. Д., Поллард, Дж. Э., Льюис, З. Т., и МакКлири, В. Р. (2009). Использование метода замены промотора показывает, что PhoU модулирует активность транспортера PstSCAB2 ABC в Escherichia coli . Заявл. Environ. Microbiol. 75, 573–582. DOI: 10.1128 / AEM.01046-1048 PubMed Аннотация | CrossRef Полный текст | Google Scholar Зальцберг, Л. И., Ботелла, Э., Hokamp, K., Antelmann, H., Maaß, S., Becher, D., et al. (2015). Полногеномный анализ связывания фосфорилированного PhoP с хромосомной ДНК выявляет несколько новых особенностей PhoPR-опосредованного ответа ограничения фосфата у Bacillus subtilis . J. Bacteriol. 197, 1492–1506. DOI: 10.1128 / JB.02570-2514 PubMed Аннотация | CrossRef Полный текст | Google Scholar Ши Л. и Хьюлетт Ф. М. (1999). Цитоплазматический киназный домен PhoR достаточен для низкой индуцируемой фосфатом экспрессии генов регулона у Bacillus subtilis . Мол. Microbiol. 31, 211–222. DOI: 10.1046 / j.1365-2958.1999.01163.x PubMed Аннотация | CrossRef Полный текст | Google Scholar Свобода, Дж. Г., Кэмпбелл, Дж., Мередит, Т. К., и Уокер, С. (2010). Стеновая функция тейхоевой кислоты, биосинтез и ингибирование. Chembiochem 11, 35–45. DOI: 10.1002 / cbic.200 PubMed Аннотация | CrossRef Полный текст | Google Scholar Tetsch, L., and Jung, K. (2009). Регуляторное взаимодействие между мембранно-интегрированными сенсорами и транспортными белками у бактерий Mol.Microbiol. 73, 982–991. DOI: 10.1111 / j.1365-2958.2009.06847.x PubMed Аннотация | CrossRef Полный текст | Google Scholar Вуппада, Р. К., Хансен, К. Р., Стрикленд, К. А. П., Келли, К. М., и МакКлири, В. Р. (2018). Передача фосфатных сигналов через альтернативные конформации транспортера фосфата PstSCAB. BMC Microbiol. 18: 8. DOI: 10.1186 / s12866-017-1126-z PubMed Аннотация | CrossRef Полный текст | Google Scholar Ван Х., Хань Х., Lv, Z., Lin, Z., Shang, Y., Xu, T., et al. (2017). PhoU2, но не PhoU1, как важный регулятор образования биопленок и устойчивости к множественным стрессам, участвуя в различных фундаментальных метаболических процессах у Staphylococcus epidermidis . J. Bacteriol. 199: e00219-17. DOI: 10.1128 / JB.00 219-17 CrossRef Полный текст | Google Scholar Weidenmaier, C., and Peschel, A. (2008). Тейхоевые кислоты и родственные гликополимеры клеточной стенки в грамположительной физиологии и взаимодействиях с хозяином. Nat. Rev. Microbiol. 6, 276–287. DOI: 10.1038 / nrmicro1861 PubMed Аннотация | CrossRef Полный текст | Google Scholar Чжэн, Дж. Дж., Синха, Д., Уэйн, К. Дж., И Винклер, М. Э. (2016). Физиологическая роль систем двойного переносчика фосфата в условиях низкого и высокого уровня фосфата и в поддержании капсулы Streptococcus pneumoniae D39. | Активация PhoPR-опосредованного ответа на ограничение фосфатов регулируется метаболизмом тейхоевой кислоты стенки у Bacillus subtilis
Введение
Активация PhoBR реагирует на перенос фосфата с помощью PstSCAB в
E. coli Варианты нормативной темы PhoBR / PstSCAB / PhoU, присутствующие в других бактериях
Активация PhoPR в
B. subtilis отвечает на метаболизм тейхоевой кислоты на стенке Варианты нормативных документов по метаболизму тейхоевой кислоты PhoPR / Wall
Заключительные замечания
Авторские взносы
Финансирование
Заявление о конфликте интересов
Благодарности
Список литературы