Какие шины установить на трактор МТЗ-82? Размер, давление в шинах, рекомендации по эксплуатации.
Трактор МТЗ-82 (Беларус) — производства Минского тракторного завода (МТЗ), является универсальным трактором класса 1,4. Основное назначение трактора МТЗ-82 — это выполнения различных сельскохозяйственных, строительных и муниципальных работ с навесным, полунавесным оборудованием и прицепами.
Колёсные трактора «Беларус» 80-ой серии, к которой относится и МТЗ-82, являются самыми массовыми тракторами не только на постсоветском пространстве, но и во всей мировой истории.
Этот трактор способен эффективно работать в самых разнообразных климатических зонах и почвенных условиях.
Типоразмер передних и задних шин трактора МТЗ-82
— передние шины: 11.2-20
11.2-20 8PR BKT TR 171 113A6 TT
11.2-20 8PR BKT TRAC FARM TT
11.2-20 8PR BKT TR 135 111A6/107A8 TT
280/85R20 (11.2R20) BKT AGRIMAX RT 855 112A8/112B TL
— задние шины: 15.
 5-38
5-38
15.5-38 8PR BKT FARM SPECIAL TT
15.5-38 8PR BKT TR 135 133A6/129A8 TT
Высокая тяга, самоочищение и устойчивость являются главными преимуществами, которые позволяют использовать шины BKT поставляемые компанией Bohnenkamp даже на самых сложных покрытиях. Шины рассчитаны на работу в различных условиях. Они характеризуются: высокой прочностью и стойкостью к проколам и порезам, длительным сроком службы.
Эксплуатация и техническое обслуживание шин для трактора МТЗ-82
Техническое состояние шин влияет на тягово-сцепные качества трактора, его расход топлива, проходимость и производительность. Таким образом, можно смело утверждать, что шины во многом определяют производительную и экономичную работу трактора. Кроме того, шины являются дорогостоящими и быстроизнашиваемыми компонентами трактора, цена которых составляет около 12% от стоимости самого трактора. В течении всего срока службы трактора шины меняются 3-4 раза, поэтому и затраты на эксплуатацию шин составляют примерно 20% общих затрат на эксплуатацию трактора.
На срок службы шины влияет не только соблюдение правил эксплуатации, но и от условий, в которых они используются: влажность и температура воздуха, особенности дорожного покрытия и свойства почвы. При эксплуатации трактора на полевых работах срок службы шин составляет около 6000 часов, а при выполнении транспортных работ они изнашиваются гораздо быстрее. На каменистых почвах, в горных районах срок службы шины сокращается на 25-35%, а на зимнем снежном покрове, наоборот, увеличивается на 25-30%.
Передние ведущие шины МТЗ-82 изнашиваются быстрее, чем задние. Это объясняется тем, что радиус качения передних шин меньше в 1,7 раза по сравнению с задними. Проходя один и тот же путь, передние шины совершают больше оборотов.
Для предотвращения преждевременного износа шин рекомендуется придерживаться следующих правил:
- строго соблюдайте нормы внутреннего давления в шинах, зависящих от условий работы и нагрузок на трактор;
 Если ведет, то остановите трактор и подкачайте шину с меньшим давлением;
Если ведет, то остановите трактор и подкачайте шину с меньшим давлением;- старайтесь не допускать резких торможений, доводящие колеса до юза, а также длительного буксования колес;
- не ставьте трактор на грунте, загрязненным нефтепродуктами;
- своевременно проверяйте и регулируйте развал-схождение передних колес;
- отключайте передний мост на сухих дорогах;
- не допускайте виляние и биение колес.
 Если ведет, то остановите трактор и подкачайте шину с меньшим давлением;
Если ведет, то остановите трактор и подкачайте шину с меньшим давлением;Давление в шинах трактора МТЗ-82
Рекомендации по внутреннему давлению в шинах, кгс/см2
| Тип работы | Шины задних колес | Шины передних колес |
|---|---|---|
| Общие виды работ | 1,4+0,1 | 1,4+0,1 |
| Для работы на мягких грунтах и пахоты | 1,0+0,1 | 1,4+0,1 |
| Для работы с тяжелым с/х оборудованием | 1,6+0,1 | 2,5+0,1 |
Нарушение норм внутреннего давлениях в шинах значительно снижает срок их эксплуатации.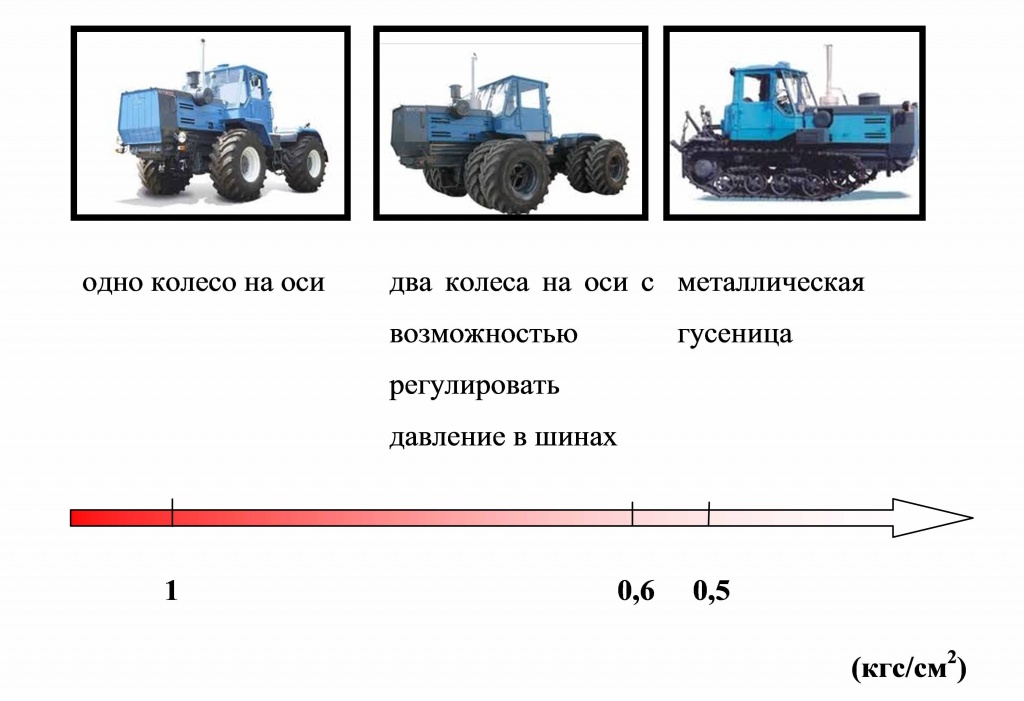 Внутреннее давление, значение которого ниже норма на 20%, сокращает срок службы на 15%, а увеличенное давление — на 10%. При выполнении определенных видов работ допустима временная перегрузка шин.
Внутреннее давление, значение которого ниже норма на 20%, сокращает срок службы на 15%, а увеличенное давление — на 10%. При выполнении определенных видов работ допустима временная перегрузка шин.
При написании статьи использованы материалы сайта tractor-mtz82.ru.
Возврат к списку
Общая техническая характеристика тракторов МТЗ-80 и МТЗ-82
Общая техническая характеристика тракторов МТЗ-80 и МТЗ-82
Трактор МТЗ-80 является новой базовой моделью семейства тракторов «Беларусь» конструкции Минского тракторного завода. Это колесный универсальный трактор с задним ведущим мостом тягового класса кН (1,4 тс). Одновременно с выпуском базовой модели начато производство ее модификации с двумя ведущими мостами — трактора МТЗ-82.
Следует отметить, что «Беларусь» наиболее распространенный, массовый колесный сельскохозяйственный трактор. Его технико-экономические и эксплуатационные показатели известны и в нашей стране и за рубежом. Однако все возрастающие требования народного хозяйства к техническому уровню тракторов поставили задачу создать вместо хорошо зарекомендовавших себя тракторов МТЗ-50 и МТЗ-52 новые, более совершенные модели МТЗ-80 и МТЗ-82. Их выпуск начат в 1974 г. Сохраняя все положительные качества своих предшественников, тракторы МТЗ-80 и МТЗ-82 заметно отличаются от них увеличенной мощностью и производительностью, лучшими условиями труда, большей надежностью и долговечностью, высоким уровнем унификации и взаимозаменяемости узлов и механизмов и более простым техническим обслуживанием.
Однако все возрастающие требования народного хозяйства к техническому уровню тракторов поставили задачу создать вместо хорошо зарекомендовавших себя тракторов МТЗ-50 и МТЗ-52 новые, более совершенные модели МТЗ-80 и МТЗ-82. Их выпуск начат в 1974 г. Сохраняя все положительные качества своих предшественников, тракторы МТЗ-80 и МТЗ-82 заметно отличаются от них увеличенной мощностью и производительностью, лучшими условиями труда, большей надежностью и долговечностью, высоким уровнем унификации и взаимозаменяемости узлов и механизмов и более простым техническим обслуживанием.
Сфера применения тракторов МТЗ-80 и МТЗ-82 чрезвычайно обширна. Они могут использоваться на самых разнообразных работах, агрегатируются с навесными, полунавесными, прицепными и стационарными машинами и орудиями. Около двухсот различных машин и орудий предназначены для работы с этими тракторами.
Рекламные предложения на основе ваших интересов:
Дополнительные материалы по теме:
Основное назначение тракторов МТЗ-80 и МТЗ-82 — комплексная механизация возделывания и уборки пропашных (кукуруза, картофель, сахарная свекла, подсолнечник, овощебахчевые) и других сельскохозяйственных культур.
Другая область применения этих тракторов — трудоемкие работы общего назначения (такие, как пахота, подготовка почвы).
Тракторы МТЗ-80, обладая высокими скоростными данными (до 35 км/ч), оснащенные пневмосистемой для торможения прицепов и светосигнальной аппаратурой, могут выполнять значительные по объему транспортные работы, успешно конкурируя, особенно в тяжелых дорожных условиях, с грузовыми автомобилями.
Приспособлены тракторы и для выполнения погрузочно-разгрузочных, дорожно-строительных и других специальных работ.
Повышенные тягово-сцепные качества и проходимость трактора МТЗ-82, оборудованного приводом на все четыре колеса и полностью сохраняющего агротехнические показатели и агрега-тируемость базовой модели, еще больше расширяют универсальность его использования и увеличивают занятость, так как позволяют применять трактор в трудных почвенных и погодных условиях как на полевых, так и на транспортных работах.
Тракторы МТЗ-80 и МТЗ-82 выполнены, как и прежние модели тракторов «Беларусь», по обычной, так называемой классической схеме (с задними колесами большего размера, чем передние) и имеют полурамную конструкцию остова.
На тракторы устанавливается дизельный двигатель Д-240 с запуском от электростартера или Д-240Л с запуском от карбюраторного пускового двигателя. (Тракторы с двигателями Д-240Л соответственно обозначаются МТЗ-80Л и МТЗ-82Л.) Увеличение мощности и улучшение топливной экономичности по сравнению с двигателями-предшественниками получено за счет использования непосредственного впрыска топлива, объемнопленочного смесеобразования в камере сгорания, перенесенной в поршень, повышения частоты вращения до 2200 об/мин и ряда других прогрессивных изменений, внесенных во все системы двигателя.
Основной и пусковой двигатели включают в работу непосредственно из кабины. Для облегчения пуска в условиях низких температур предусмотрен электрофакельный подогреватель всасывающего коллектора и съемный жидкостной подогреватель для водяной и масляной систем двигателя.
Силовая передача (трансмиссия) обеспечивает расширенный и рациональный ряд скоростей. Число передач увеличено до восемнадцати, что дает возможность эффективно загрузить двигатель в рабочем диапазоне передач, в том числе на повышенных скоростях (9—15 км/ч), а также в области транспортных передач, где скоростной ряд также увеличен (17,9, 25,3 и 33,4 км/ч). Хо-доуменьшитель, который при необходимости устанавливают на левой стороне коробки передач, позволяет получить дополнительные пониженные технологические скорости для специальных работ (до 0,26 км/ч).
Хо-доуменьшитель, который при необходимости устанавливают на левой стороне коробки передач, позволяет получить дополнительные пониженные технологические скорости для специальных работ (до 0,26 км/ч).
Муфта сцепления снабжена демпферными пружинами — гасителями крутильных колебаний коленчатого вала двигателя. На силовом валу муфты установлен дисковый тормозок, который останавливает вал в конце выключения муфты и тем самым способствует облегченному переключению и безударному введению в зацепление шестерен коробки передач.
Дифференциал заднего моста блокируется фрикционной муфтой с гидравлическим приводом, связанным с управлением гидроусилителем рулевого управления.
Рис. 1. Трактор МТЗ-80
Рис. 2. Трактор МТЗ-82
Независимый и синхронный привод заднего вала отбора мощности выполнен двухскоростным, боковой вал отбора мощности и приводной шкив также имеют две передачи.
В привод переднего ведущего моста трактора МТЗ-82 введена фрикционная предохранительная муфта, которая монтируется в промежуточной опоре карданной передачи.
С целью обеспечения устойчивости и управляемости трактора, а также повышения эффективности действия переднего ведущего моста при навеске машин, разгружающих передние колеса, на кронштейн переднего бруса устанавливают комплект легкосъемных балластных грузов.
Для повышения тягово-сцепных качеств и проходимости в тяжелых почвенных условиях предусмотрена установка резинометаллического полугусеничного хода.
Раздельно-агрегатная гидросистема обеспечивает высотный и дополнительно силовой и позиционный способы регулирования положения навесных почвообрабатывающих машин относительно трактора. Силовое и позиционное регулирование — автоматическое, на основе датчиков, которые встроены в механизм навески трактора и реагируют на изменения тягового сопротивления и положения орудия.
Чтобы повысить эффективность применения на транспортных работах, тракторы оборудованы универсальной пневматической системой, которая обеспечивает привод тормозов большегрузных транспортных прицепов и других машин. Пневмосистему можно использовать также для накачки шин и некоторых других целей.
Кабина трактора установлена на резиновых амортизаторах и не имеет жесткой связи с остовом трактора, благодаря чему снижается уровень вибрации и шума. Для обивки дверей и стенок кабины используются теплошумоизоляционные материалы, применяемые в современном автомобилестроении. Обогревается кабина отопителем, который отбирает тепло от водяной системы двигателя. В условиях запыленности и в жару в кабину подается предварительно очищенный и увлажненный воздух. Естественная вентиляция кабины через открывающиеся крышу и заднее окно.
Одноместное сиденье тракториста снабжено торсионной подвеской и гидравлическим амортизатором, его положение регулируется в зависимости от веса и роста водителя. Положение рулевого колеса также может быть изменено для удобства управления.
Жесткость каркаса кабины отвечает требованиям безопасности при аварийном опрокидывании трактора. Форма облицовки кабины и в целом трактора рационально увязана с удобством обслуживания и условиями труда.
Благодаря применению прогрессивных решений в конструкции узлов и механизмов значительно снижена трудоемкость технических обслуживаний, сокращено число точек смазки, увеличено большинство межрегулировочных сроков, часть регулировок снята, упрощена или проводится только после выработки моторесурса (при ремонтах). Многие операции по обслуживанию и управлению облегчены, механизированы и автоматизированы.
Повышение энергонасыщенности тракторов МТЗ-80 и МТЗ-82, улучшение условий труда, оборудование тракторов автоматической блокировкой дифференциала, гидросистемой с силовым и позиционным регулированием, пневмосистемой и ряд других прогрессивных изменений конструкции позволяют значительно повысить их производительность (по сравнению с тракторами МТЗ-50 и МТЗ-52 в среднем на 35%).
все тот же «Беларус» в новом обличии
02.07.2013
Трактор МТЗ 82.1: все тот же «Беларус» в новом обличииСиние, шустрые и неприхотливые — тракторы МТЗ 80-й серии заняли видное место во многих сферах человеческой деятельности и стали, без преувеличения, легендой отечественного тракторостроения. Трактор МТЗ-82.1 — одна из самых современных модификаций знакомого «Беларуса», и именно эта машина стала главным героем этой статьи.
История трактора МТЗ-82.1
Первый серийный трактор МТЗ-82.1 сошел с конвейера Минского тракторного завода в 2000-х годах, однако история этой машины началась гораздо раньше — фактически, с середины прошлого века. Так, еще в 1957 году был запущен в производство универсальный трактор МТЗ-5, с 1962 года на производство поставлены тракторы МТЗ-50 и МТЗ-52, ставшие глубокой модернизацией трактора МТЗ-5, а в 1974 году свет увидели первые тракторы МТЗ-80 и МТЗ-82, которые, в свою очередь, стали глубокой модернизацией машин МТЗ-50 и МТЗ-52 соответственно. Именно поэтому даже сейчас можно найти унифицированные запчасти на МТЗ-52 и МТЗ-82.1, хотя эти машины и выпущены в разное время в разных странах.
Именно поэтому даже сейчас можно найти унифицированные запчасти на МТЗ-52 и МТЗ-82.1, хотя эти машины и выпущены в разное время в разных странах.
Тракторы МТЗ серии 80 выпускаются и по сей день — такое «долгожительство» объясняется тем, что эти машины получились именно такими, какие нужны фермерам, строителям, дорожным службам, лесным хозяйствам, службам ЖКХ и всем, кому просто нужна мощность, неприхотливость и универсальность.
Интересно, что тракторы МТЗ-80 и МТЗ-82 с 2000 годов выпускаются не под аббревиатурой МТЗ, а под названием «Беларус», поэтому не нужно удивляться, что наш герой — МТЗ-82.1 — поставляется с Минского тракторного завода под наименованием «Беларус-82.1».
Технические характеристики и особенности
МТЗ-82.1 — универсальный трактор тягового класса 1,4, имеет традиционную компоновку и конструкцию, которая практически не претерпела изменений за последние полвека. Трактор МТЗ-82.1, в отличие от «младшего» брата МТЗ-80.1, имеет колесную формулу 4×4, а значит, повышенную проходимость, что позволяет использовать его в самых сложных дорожных и климатических условиях. Также в машине предусмотрена возможность сдваивания задних колес.
Также в машине предусмотрена возможность сдваивания задних колес.
Главное отличие модификации 82.1 от базовой модели МТЗ-82 заключается в кабине: у современной модификации она большая и комфортабельная, обеспечивает широкий обзор и создает комфортные условия работы. Также новая кабина соответствует повышенным требованиям к безопасности водителя, что открыло путь трактору на европейский рынок.
Что касается силовой установки, то за последние десятилетия практически ничего не изменилось: на трактор МТЗ-82.1 устанавливается все тот же Д-243 Минского моторного завода мощностью 80-82 л.с., хотя многие современные модификации трактора оборудованы электрическим стартером. Использование одной марки двигателя на разных модификациях тракторов легко решает проблему ремонта — запчасти на МТЗ-82.1 в большинстве своем унифицированы с предыдущими модификациями машины, что решает проблему их поиска и покупки.
Благодаря продуманной и надежной гидравлической системе трактор может работать с самым широким спектром навесного, полунавесного и прицепного оборудования различного назначения. Собственно, эта способность и является одним из главных преимуществ тракторов МТЗ.
Собственно, эта способность и является одним из главных преимуществ тракторов МТЗ.
Недостатки трактора МТЗ-82.1
Тракторы МТЗ уже несколько десятилетий верой и правдой служат людям, и в целом отношение к ним положительное, однако современные модификации нередко вызывают недовольство у потребителей. Главная причина — мнение об общем снижении качества производства узлов и агрегатов, и сборки трактора после развала СССР. Трудно судить о том, насколько эти нарекания объективны, ведь очень многое зависит от условий эксплуатации и содержания трактора — у плохого хозяина даже новенький МТЗ-82.1 быстро выйдет из строя.
Но в любом случае, тракторы МТЗ нуждаются в периодическом ремонте, поэтому запчасти на МТЗ-82.1 и другие старые и новые модификации пользуются стабильным спросом на рынке, а благодаря их доступности позволяют решать проблему ремонта «рабочих лошадок» с минимальными затратами.
Другие статьи
#Стойка стабилизатора Nissan
Стойка стабилизатора Nissan: основа поперечной устойчивости «японцев»
22. 06.2022 | Статьи о запасных частях
06.2022 | Статьи о запасных частях
Ходовая часть многих японских автомобилей Nissan оснащается стабилизатором поперечной устойчивости раздельного типа, соединенным с деталями подвески двумя отдельными стойками (тягами). Все о стойках стабилизатора Nissan, их типах и конструкции, а также о подборе и ремонте — читайте в данной статье.
#Ремень приводной клиновой
Ремень приводной клиновой: надежный привод агрегатов и оборудования
15.06.2022 | Статьи о запасных частях
Для привода агрегатов двигателя и в трансмиссиях различного оборудования широко применяются передачи на основе резиновых клиновых ремней. Все о приводных клиновых ремнях, их существующих типах, особенностях конструкции и характеристиках, а также о правильном выборе и замене ремней — читайте в статье.
Барабан тормозной ГАЗ: управляемость и безопасность горьковских автомобилей
08.06.2022 | Статьи о запасных частях
Тормозные системы большинства ранних и актуальных моделей автомобилей ГАЗ оснащаются колесными механизмами барабанного типа. Все о тормозных барабанах ГАЗ, их существующих типах, конструктивных особенностях и характеристиках, а также о выборе, замене и обслуживании данных деталей — читайте в статье.
Все о тормозных барабанах ГАЗ, их существующих типах, конструктивных особенностях и характеристиках, а также о выборе, замене и обслуживании данных деталей — читайте в статье.
#Палец поршневой
Палец поршневой: прочная связь поршня и шатуна
02.02.2022 | Статьи о запасных частях
В любом поршневом двигателе внутреннего сгорания присутствует деталь, соединяющая поршень с верхней головкой шатуна — поршневой палец. Все о поршневых пальцах, их конструктивных особенностях и способах установки, а также о верном подборе и замене пальцев различных типов подробно рассказано в статье.
Вернуться к списку статей
5.3. Проходимость
Проходимость трактора (автомобиля) — одно из основных свойств, определяющих эксплуатационную надежность.
Проходимость
зависит в основном от удельного давления
на
почву, тягово-сцепных свойств колес,
дорожного просвета, шрины
колеи, а также размеров защитных зон и
агротехни ческого
просвета (при междурядной обработке
пропашных культур).
Удельное давление на почву — это часть веса трактора (Н), приходящаяся на 1 см2 опорной поверхности колеса. Оно зависит от нагрузки на колесо, давления воздуха в шине, размеров шин, жесткости и степени погружения колес в почву. Для тракторов, работающих на мягких почвах, давление в передних колесах целесообразно снижать до 0,08…0,11 МПа, а на транспортных работах, при сравнительно плотной дороге, увеличивать до 0,14…0,25 МПа.
Тягово-сцепные свойства колес зависят в основном от состояния почвы, сцепного веса машины, рисунка на протекторе колес и его износа. При износе протектора колес более 80 % возрастает их буксование, что приводит к снижению производительности тракторного агрегата и повышению расхода топлива. Повышение тягово-сцепных свойств машин достигается изменением давления воздуха в шинах, наполнением камер ведущих колес водой,
установкой
сдвоенных шин, дополнительных грузов
на передний брус и диски колес и т. д.
д.
Дорожный просветh — это расстояние в миллиметрах от поверхности почвы (дороги) до нижних точек трактора (автомобиля), расположенных обычно на переднем или заднем мосту (рис. 5.10). Дорожный просвет тракторов общего назначения составляет 300…400 мм, а универсально-пропашных — 400… 830 мм и более (например, у тракторов с портальным остовом).
Рис. 5.10. Схема колесных универсальных и пропашных тракторов:
а — четырехколесный универсально-пропашной; б — пропашной со сближенными передними колесами; в — пропашной с одинарным передним колесом; г —пропашной с портальным остовом; д — схема (абрис) впи-сываемости колесного трактора в междурядья
Агротехнический просвет —это просвет между трактором и рядками растений в момент их обработки.
Ширина колеи — расстояние между осевыми
линиями, проведенными через середины
профилей шин (гусениц). Универсально-пропашные
тракторы имеют регулируемую ширину
колеи. Например, у трактора МТЗ-80 и его
модификаций колея передних колес
регулируется в пределах 1200… 1800 мм, а
задних — 1400…2100 мм.
Например, у трактора МТЗ-80 и его
модификаций колея передних колес
регулируется в пределах 1200… 1800 мм, а
задних — 1400…2100 мм.
Проходимость трактора в междурядьях пропашных культур характеризуется защитной зоной (х — внутренней, у — наружной), представляющей собой расстояние от середины рядка до ближайших точек ходовой части трактора.
Размеры защитных зон (см. рис. 5.10, д)
где b — ширина междурядий, мм; я — число рядков растений под трактором; В -ширина колеи, мм; с — ширина покрышки заднего колеса трактора, мм.
Глава 6
РАБОЧЕЕ И ВСПОМОГАТЕЛЬНОЕ ОБОРУДОВАНИЕ ТРАКТОРОВ И АВТОМОБИЛЕЙ
6.1. Рабочее оборудование
С целью полной реализации потенциальных возможностей и показателей мощности тракторы снабжены различным рабочим оборудованием.
На современных тракторах используют
гидронавесную систему, регулятор глубины
обработки почвы, догружатель ведущих
колес, вал отбора мощности, приводной
шкив, прицепное устройство. К рабочему
оборудованию автомобилей относят
прицепное устройство, лебедку,
приспособление для накачивания шин,
различные приборы.
К рабочему
оборудованию автомобилей относят
прицепное устройство, лебедку,
приспособление для накачивания шин,
различные приборы.
Гидравлическая навесная система служит для соединения навесных машин и орудий с трактором, а также перевода их в рабочее и транспортное положение. Она состоит из навесного устройства и гидравлического привода (системы). Навесное устройство служит для соединения трактора с навесными машинами.
Трактор, гидравлическая навесная система и машина образуют навесной агрегат. Навесные агрегаты обладают существенными преимуществами перед прицепными: хорошая маневренность, более высокая производительность, меньший расход топлива на единицу выполненной работы, относительно малая металлоемкость навесных машин. Кроме того, на некоторых
видах работ не нужен вспомогательный обслуживающий персонал.
Возможны различные варианты размещения
навесных машин в тракторном агрегате
(рис. 6.1): задняя, передняя, фронтальная,
боковая, эшелонированная, шеренговая,
комбинированная. В комбинированных
агрегатах, когда одновременно совмещаются
несколько технологических операций
(например, культивация, посев и подкормка
минеральными удобрениями), применяют
одновременно два варианта навески,
например фронтальную и заднюю.
В комбинированных
агрегатах, когда одновременно совмещаются
несколько технологических операций
(например, культивация, посев и подкормка
минеральными удобрениями), применяют
одновременно два варианта навески,
например фронтальную и заднюю.
Навесная система для присоединения сельскохозяйственных машин сзади состоит из гидроцилиндра 1 (рис. 6.2, а), вала с рычагом, двух подвесных рычагов, соединенных раскосами с нижними продольными тягами, центральной тяги. Рычаг 2 вала 3 соединен со штоком гидроцилиндра /. Передние концы продольных тяг 8 в точках А’, В и центральной шарнирно присоединены к корпусу трансмиссии трактора, а их задние концы в точках А, Б, В к оси подвеса в точках А, Б рабочей машины или орудия (рис. 6.2, б) и к стойке в точке В. Различают двух- и трехточечные навесные устройства.
Рис. 6.2. Устройство механизма навески:
а — на тракторе; б—на машине или орудии; 1 — гидроцилиндр; 2 — рычаг; 3 — вал; 4 — правый и левый рычаги; 5 — правый и левый раскосы; 6— центральная тяга; 7— стяжная гайка; 8— левая и правая продольные тяги; А, Б, В, Г, А’, Б’, В’— точки присоединения
У двухточечного навесного устройства
обе продольные тяги соединены в точке
Г. Таким образом, продольные и центральная
тяги имеют две точки крепления к корпусу
трансмиссии трактора. У трехточечного
(рис. 6.2, а) навесного устройства продольные
тяги крепятся раздельно в точках Аи Б
и таким образом продольные и центральная
тяги имеют три точки крепления к корпусу
трансмиссии. На гусеничных тракторах
предусматривается переоборудование
механизма навески из двухточечной в
трехточечную и наоборот. Колесные
тракторы оснащены трехточечным навесным
устройством.
Таким образом, продольные и центральная
тяги имеют две точки крепления к корпусу
трансмиссии трактора. У трехточечного
(рис. 6.2, а) навесного устройства продольные
тяги крепятся раздельно в точках Аи Б
и таким образом продольные и центральная
тяги имеют три точки крепления к корпусу
трансмиссии. На гусеничных тракторах
предусматривается переоборудование
механизма навески из двухточечной в
трехточечную и наоборот. Колесные
тракторы оснащены трехточечным навесным
устройством.
Рассмотрим устройство и действие отдельных узлов гидравлической системы на примере гидросистемы трактора МТЗ-80 и
его модификаций (рис. б.З).
В гидравлическую систему входят
шестеренный насос НШ-32-2 (НШ — насос
шестеренный, цифры — теоретическая
подача жидкости в см3 на один оборот вала привода насоса),
основной Ц-100 и два выносных Ц-75 цилиндра
(Ц — цилиндр, цифры — внутренний диаметр
корпуса в миллиметрах), распределитель
Р75-33-Р, силовой (позиционный) регулятор,
гидравлический увеличитель сцепного
веса (ГСВ), гидроаккумулятор, корпус
гидроагрегатов с фильтром и шланги
высокого давления с запорным устройством.
Насос 1 через всасывающий патрубок 2 забирает масло из бака и под давлением более 10 МПа подает по маслопроводу к распределителю 6 и силовому регулятору 13. Распределитель регулирует направление потока масла. Он направляет масло либо в бак по сливному маслопроводу, пропуская его через фильтр, либо по промежуточному маслопроводу в ГСВ. Далее по маслопроводу масло поступает в силовой регулятор и по рукаву высокого давления в гидроцилиндр или через боковые выводы непосредственно к гидроприводу сельскохозяйственных машин.
Неподвижно закрепленные на тракторе устройства гидросистемы соединяют стальными бесшовными трубопроводами, рассчитанными на давление до 32 МПа, а гидроцилиндрам жидкость подводится по гибким шлангам. Маслопроводы соединяют с помощью специальных муфт, снабженных самозапирающимися устройствами шарового типа.
Регулятор глубины обработки почвы работает следующим образом.
Верхняя центральная тяга навесного
устройства соединена с корпусом заднего
моста трактора не жестко, как обычно, а
болтом через пластинчатую пружину. При
заглублении машины, например плуга,
сверх нормы увеличивается давление на
пружину, в результате чего ее длина
уменьшается, а поводок через тягу и
рычаг силового регулирования перемещает
золотник силового регулятора вверх, в
результате чего масло направляется в
гидроцилиндр и плуг выглубляется.
При
заглублении машины, например плуга,
сверх нормы увеличивается давление на
пружину, в результате чего ее длина
уменьшается, а поводок через тягу и
рычаг силового регулирования перемещает
золотник силового регулятора вверх, в
результате чего масло направляется в
гидроцилиндр и плуг выглубляется.
Как только глубина обработки почвы достигнет заданной величины, уменьшится воздействие на пружину, она удлинится, возвратит золотник регулятора в исходное положение и подача масла в цилиндр прекратится. Включение (и выключение) регулятора в систему осуществляется рычагом переключателя.
Если навешенные на трактор
машина или орудие удерживаются во время
работы в заданном положении (позиции)
относительно остова трактора независимо
от тягового сопротивления, например
при посеве на поле с ровным рельефом,
то золотник регулятора соединяется
через тягу с поворотным рычагом,
посредством которого шток гидроцилиндра
соединен с навесным устройством. При
перемещении рычага сигнал через тягу
передается на золотник силового
регулятора, который для подъема или
опускания рабочей машины направляет
масло в гидроцилиндр.
Догружатель ведущих колес бывает двух типов: механический, когда сцепной вес увеличивают за счет веса агрегатируемои машины, перенося переднюю точку присоединения центральной тяги (чем ниже точки присоединения тяги, тем больше сцепной вес), и гидравлический (ГСВ).
Гидравлический догружатель или увеличитель сцепного веса расположен на стенке корпуса гидроагрегатов справа от распределителя. Работает он следующим образом. При недостаточном сцепном весе тракторного агрегата (ведущие колеса начинают пробуксовывать) с помощью ГСВ в гидроцилиндр под небольшим давлением (0,8…0,35 МПа) подается масло. При этом навесное устройство стремится поднять навешенную машину в транспортное положение, но давления, создающего подъемную силу 300…500 Н, для этого недостаточно. Тем не менее усилие передается через навесное устройство на корпус трактора, прижимая его задние колеса к почве и уменьшая их буксование.
Вал
отбора мощности (ВОМ) предназначен для
привода рабочих органов, агрегатируемых
с тракторами передвижных или стационарных
машин. По месту расположения ВОМ может
быть задний, боковой (рис. 6.4) и передний.
Наиболее распространены задние ВОМ —
их имеют все тракторы, за исключением
самоходного шасси Т-16М.
По месту расположения ВОМ может
быть задний, боковой (рис. 6.4) и передний.
Наиболее распространены задние ВОМ —
их имеют все тракторы, за исключением
самоходного шасси Т-16М.
Рис. 6.4. Валы отбора мощности:
а а— положение на тракторе; б — вид сзади; / — боковой ВОМ; 2—задний ВОМ
Универсальные тракторы (МТЗ-80, Т-40М и др.), кроме заднего, оборудованы боковым ВОМ. Все агрегатируемые с самоходным шасси Т-16М машины размещают на специальной раме нмгреди двигателя, поэтому здесь применяют передний ВОМ.
По характеру привода
различают зависимый, независимый и
ппхронный ВОМ. Если ВОМ приводится во
вращение от одно-ГО КЗ валов трансмиссии,
то его работа зависит от включения и
iвключения
муфты сцепления трактора: при выключении
муфты мсмления вместе с остановкой
трактора прекращается вращение in
>М. Привод ВОМ такого типа называется
зависимым. Независиимый ВОМ получает
вращение от специального вала, соединен
с двигателем через отдельную муфту
сцепления или двухпо-тчную муфту, а
иногда через планетарный механизм, что
позволяет выключать ВОМ независимо от
выключения главного сцепления трактора.
Независимый и зависимый ВОМ имеет две стандартные части, вращения —540 и 1000 мин»1 при номинальной частоте [мщения коленчатого вала двигателя.
Синхронный ВОМ приводится во вращение от вала, соединенного постоянной передачей с вторичным валом коробки передач. Поэтому частота его вращения изменяется с переменой передачи, но остается постоянной на 1 м пути (3,5 мин»1). Такой КОМ необходим при посеве, работе с разбрасывателями удобрении и т.д.
Приводной шкивприменяют для передачи момента вращения помощью ременной передачи от трактора рабочим органам машины при использовании трактора в стационарном положении. Шкив устанавливают в задней части трактора. Он приводится во вращение от ВОМ.
Прицепное устройство служит для буксировки прицепных машин
и тележек (прицепов). Оно состоит из
скобы (поперечины), Закрепленной в
кронштейнах остова трактора, и серьги,
присоединенной к скобе пальцами. Устройство размещают сзади трак-юра.
Оно позволяет регулировать точку
присоединения машин и тележек к трактору
и горизонтальной плоскости, I
v
большинства тракторов и но высоте.
Устройство размещают сзади трак-юра.
Оно позволяет регулировать точку
присоединения машин и тележек к трактору
и горизонтальной плоскости, I
v
большинства тракторов и но высоте.
Техническое описание тракторов Беларус 80.1 и 82.1. Характеристики трактора МТЗ 80 и 82.
Область применения тракторов Беларус 80.1 и 82.1
Колесные тракторы Беларус 80-й серии являются универсально-пропашными тракторами, предназначенными как для возделывания различных культур (например, корнеплодов), так и для разнообразных работ в сельском, коммунальном и лесном хозяйствах, на транспорте, в строительстве и прочих отраслях.
Тракторы относятся к тяговому классу 1,4, что соответствует развиваемому ими номинальному тяговому усилию 14 кН.
Конструкционная масса трактора МТЗ:
-
Беларус 82.1 – 3750 кг,
-
Беларус 80.1 – 3520 кг.
Рассмотрим подробнее некоторые особенности конструкции этих машин.
Остов и ходовая система
Конструкция тракторов МТЗ 80 – полурамная, как и у большинства тракторов Минского тракторного завода. Передняя полурама, состоящая из литого переднего бруса и продольных штампованных лонжеронов, совместно с несущими картерами корпуса муфты сцепления, коробки передач и заднего моста образует прочный остов трактора. Двигатель передними опорами через резиновые амортизаторы эластично установлен на переднем брусе, задняя же часть двигателя картером маховика жестко соединена с корпусом муфты сцепления.
Ходовая система тракторов снабжена пневматическими шинами низкого давления. Тракторы имеют передние управляемые колеса, при этом ведущие колеса Беларус 82.1 – передние и задние (колесная формула 4К4), а ведущие колеса Беларус 80.1 – только задние (4К2). Для уменьшения давления на грунт и улучшения проходимости тракторов конструкцией предусмотрена возможность сдваивания задних колес.
Для улучшения сцепных качеств при работе с различным сельскохозяйственным оборудованием возможна установка дополнительных грузов на передний брус или на диски задних колес.
Двигатель
На тракторах устанавливается четырехцилиндровый безнаддувный (атмосферный) дизельный двигатель Д-243 номинальной мощностью 59,6 кВт (81 л.с.) при 2200 об/мин производства Минского моторного завода, рабочий объем двигателя – 4,75 л.
Двигатель имеет комбинированную систему смазки — смазка деталей производится под давлением либо разбрызгиванием. Система смазки включает в себя масляный картер двигателя, шестеренный масляный насос, радиатор охлаждения и полнопоточный масляный фильтр с предохранительным клапаном.
Топливная система двигателя включает в себя топливный насос низкого давления (подкачивающий насос), топливный насос высокого давления (ТНВД), фильтры грубой и тонкой очистки топлива и топливопроводы низкого и высокого давления.
Двигатель снабжен электростартерной системой пуска. Для облегчения пуска при низких температурах применяется электрофакельная система пускового подогрева, установленная на впускном коллекторе.
Основным элементом системы питания двигателя воздухом является воздушный фильтр с двухступенчатой очисткой воздуха — сухой центробежной и масляной инерционно-контактной.
Охлаждение двигателя осуществляется принудительной циркуляцией охлаждающей жидкости от центробежного насоса, который, в свою очередь, приводится во вращение клиновым ремнем от шкива коленчатого вала. Жидкость охлаждается при прохождении через радиатор, расположенный в передней части трактора перед двигателем. Радиатор охлаждается потоком воздуха от постоянно работающего лопастного вентилятора.
Ускорение прогрева двигателя после пуска, а также установка оптимального температурного режима достигается за счет термостата, регулирующего поток охлаждающей жидкости.
Трансмиссия
Тракторы имеют фрикционную однодисковую постоянно замкнутую муфту сцепления с механическим управлением. Выключение муфты сцепления производится педалью. Ведомый диск муфты имеет безасбестовые накладки, по заказу может устанавливаться ведомый диск с износостойкими металлокерамическими накладками.
Коробка передач – механическая, 9-ступенчатая, двухдиапазонная, с двухступенчатым понижающим редуктором, не синхронизированная, обеспечивает движение на 18 передачах переднего и 4 передачах заднего хода. Максимальная расчетная скорость движения тракторов – 34,3 км/ч, минимальная – 1,94 км/ч. Конструкция коробки передач предусматривает возможность установки гидравлического ходоуменьшителя.
Задний мост – с главной конической передачей, блокируемым дифференциалом и цилиндрическими конечными передачами.
Передний мост тракторов Беларус 80-й серии – портального типа. Ведущий передний мост трактора Беларус 82.1 включает в себя главную коническую передачу, самоблокирующийся дифференциал и конечные передачи — двухступенчатые конические колесные редукторы. На некоторые модификации (например, Беларус 82.1-23/12-23/32, Беларус 820) устанавливается передний мост повышенной несущей способности с планетарно-цилиндрическими редукторами. Привод переднего ведущего моста осуществляется от раздаточной коробки двумя карданными валами с промежуточной опорой со встроенной предохранительной муфтой. Предусмотрены три режима работы переднего ведущего моста: выключен, включен принудительно и автоматически включен при буксовании задних колес.
Предусмотрены три режима работы переднего ведущего моста: выключен, включен принудительно и автоматически включен при буксовании задних колес.
Передний мост (передняя ось) трактора Беларус 80.1 имеет возможность бесступенчатого изменения колеи передних колес благодаря применению выдвижных кулаков, на которых установлены цапфы с колесными полуосями.
Тормозная система
Рабочие тормоза – дисковые, сухие, установленные на валах ведущих шестерен конечных передач задних колес. Управление тормозами – механическое, от педалей, раздельное для правого и левого бортов. Предусмотрена блокировка педалей для транспортного режима движения. Стояночный тормоз – дисковый, сухой, с независимым ручным управлением.
Для управления тормозами прицепа тракторы снабжены пневмосистемой, питание которой осуществляется от компрессора, установленного на двигателе. Пневмопривод управления тормозами прицепа – однопроводный, сблокированный с управлением тормозами трактора. Давление в пневмосистеме поддерживается регулятором в пределах 0,65-0,8 МПа. Максимальная масса буксируемого прицепа при сблокированных тормозах прицепа и трактора – 9000 кг.
Давление в пневмосистеме поддерживается регулятором в пределах 0,65-0,8 МПа. Максимальная масса буксируемого прицепа при сблокированных тормозах прицепа и трактора – 9000 кг.
Вал отбора мощности
Для привода рабочего оборудования тракторы имеют задний вал отбора мощности, работающий в независимом двухскоростном режиме – 540 об/мин (при скорости двигателя 1632 об/мин) и 1000 об/мин (при скорости двигателя 1673 об/мин) или синхронном режиме — 3,44 оборота на метр пути). Направление вращения вала – по часовой стрелке. В стандартную комплектацию тракторов входит хвостовик вала отбора мощности на 8 зубьев для скорости 540 об/мин, для скорости 1000 об/мин необходимо установить хвостовик на 21 зуб.
Рулевое управление
На тракторы устанавливается гидрообъемное рулевое управление. Питание гидросистемы рулевого управления производится шестеренным гидронасосом левого вращения типа НШ-10. Применяется насос-дозатор героторного типа, без реакции на рулевом колесе. Механизм поворота колес – исполнительный гидроцилиндр двойного действия и рулевая трапеция.
Механизм поворота колес – исполнительный гидроцилиндр двойного действия и рулевая трапеция.
Наименьший радиус поворота трактора – 4,5 м (с подтормаживанием заднего внутреннего колеса).
На тракторы ранних выпусков устанавливалось гидромеханическое рулевое управление с гидроусилителем.
Гидронавесная система
Рабочая гидросистема тракторов Беларус – раздельно-агрегатная. Подача гидрожидкости от гидробака осуществляется шестеренным насосом НШ-32 правого вращения с объемной подачей не менее 45 л/мин при 2200 об/мин вала двигателя. Давление срабатывания предохранительного клапана гидросистемы – 18-20 МПа.
В основном, тракторы Беларус 80.1 и Беларус 82.1 имеют стандартную гидронавесную систему без силового регулятора, которая обеспечивает только высотное регулирование положения орудий. Такая система имеет две пары независимых гидровыводов справа и слева и одну дублированную пару в задней части трактора. Однако, на некоторые машины устанавливается гидронавесная система с силовым позиционным регулятором, позволяющая производить автоматическое регулирование как в высотном, так и в силовом и позиционном режимах, что позволяет повысить качество обработки почвы и улучшить условия труда тракториста. В такой системе имеется три пары независимых гидровыводов.
В такой системе имеется три пары независимых гидровыводов.
Заднее навесное устройство
На тракторы устанавливается заднее навесное устройство трехточечного типа, категории НУ-2, с внутренней или наружной блокировкой нижних тяг. Грузоподъемность заднего навесного устройства на оси подвеса – не менее 3200 кг.
Аренда трактора беларус 82.1 в Москве и области по доступным ценам
Заказать технику
Дополнительное оборудование: отвал, щетка, цистерна, прицеп
12 000 ₽ / смена
7+1 ч.
, последующий час 1 800 ₽
 , последующий час 1 800 ₽
, последующий час 1 800 ₽| Технические характеристики | Значения |
| Длина | 3930 |
| Ширина | 1970 |
| Высота | 2800 |
| Максимальная ширина захвата щётки, мм | 1800 |
| Эксплуатационная масса | 4 тонны |
Аренда трактора беларус 82.1
Трактор МТЗ Белорусского производства пользуется огромной популярностью в нашей стране. Особенно популярна модель, которая носит название Беларус 82,1. Это самая распространенная машина на сегодняшний день. За время многолетнего использования техника зарекомендовала себя как универсальная, простая в обслуживании и доступная по цене. Эти составляющие сделали данный трактор самым успешным в своем сегменте.
Беларус 82,1 это последняя модификация модели 82, которая была поставлена на конвейер в далеком 1972 году. Техническая составляющая осталась прежней, но большинство узлов и механизмов в последние годы усовершенствовались, что позволило эффективно эксплуатировать этот трактор в современных условиях.
Техническая составляющая осталась прежней, но большинство узлов и механизмов в последние годы усовершенствовались, что позволило эффективно эксплуатировать этот трактор в современных условиях.
Аренда трактора беларусь имеет множество преимуществ. Но для начала давайте рассмотрим его конструкционные и технические особенности:
- машина оснащена дизельным двигателем объемом 4,75 л и мощностью 81 л.с.;
- объем топливного бака составляет 130 л, что дает возможность продолжительное время функционировать без дозаправки;
- данная модель имеет механическую коробку передач;
- мтз 82,1 полноприводный. Имеет функцию блокировки дифференциала заднего моста;
- максимальная скорость передвижения 34 км/ч;
- тормозная система оснащена пневматическим приводом;
- гидравлическая система имеет три пары выводов, что позволяет оператору управлять навесным оборудованием;
- трактор оснащен двумя валами отбора мощности, которые приводят в действие навесное оборудование.

Аренда трактора мтз 82 — это отличный выбор для решения многочисленных задач. На него можно дополнительно устанавливать навесное оборудование, делая сферу применения машины очень разнообразной:
- строительство. Беларус 82,1 можно эксплуатировать в качестве экскаватора, бульдозера и ямокопателя;
- аграрный сектор. Трактор успешно применяется для подготовки и обработки почвы, сбора и транспортировки сельхоз продукции. Это возможно благодаря установке прицепного оборудования, плугов, сеялок и т.д.;
- лесное хозяйство. Имея высокую проходимость и хорошую тяговую силу, эта техника подходит для транспортировки леса-кругляка. Также трактор можно использовать как лесоукладчик;
- коммунальное хозяйство. МТЗ 82 может выполнять функции тягача, погрузчика, очистной и снегоуборочной машины. Помимо этого, он используется для проведения ремонтных работ водопроводов и канализации;
- перевозка грузов. Отличные ходовые качества и высокая проходимость позволяют трактору транспортировать груз в условиях бездорожья.
 Отличные ходовые качества и высокая проходимость позволяют трактору транспортировать груз в условиях бездорожья.
Отличные ходовые качества и высокая проходимость позволяют трактору транспортировать груз в условиях бездорожья.Трактор погрузчик беларус мтз 82.1 аренда
Аренда трактора мтз широко востребована из-за его экономичности, универсальности, высокой проходимости и возможности использования широкого спектра навесного оборудования.
Имея простую и надежную конструкцию, которая остается неизменной уже несколько десятилетий, трактор не испытывает затруднений с ремонтом. К нему легко найти как новые запчасти, так и бывшие в употреблении. Это позволяет значительно снизить расходы на проведение ремонта, что является весомым преимуществом по сравнению с зарубежными тракторами.
Трактор погрузчик беларус мтз 82.1 аренда — новая услуга, которая доступна в нашей компании с недавнего времени. Благодаря качественному навесному оборудованию, которые мы приобретаем непосредственно у производителя, наши клиенты теперь имеют возможность использовать трактор в качестве погрузчика на различных производственных объектах.
Трактор мтз 82.1 заказать проще простого. Достаточно позвонить нам по телефону указанному на сайте и наш консультант предоставит вам полную информацию по условиям составления договора аренды. Техника будет доставлена на объект в течение кратчайшего времени.
Модуль Boresight — Martin Marietta Corporation
СВЯЗЬ С ДРУГИМ ИЗОБРЕТЕНИЕМ
Это изобретение может рассматриваться как тесно связанное с одновременно находящейся на рассмотрении патентной заявкой Макса Амона и Андре Массона под названием «Command Optics», сер. № 571,581, поданной 17 января 1984 г.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Автоматические системы телевизионного слежения, включающие телевизионные точечные устройства слежения или средства слежения за площадной корреляцией, работающие с совместимыми датчиками, такими как видиконы, оказались способными удовлетворить требования к точности системного наведения на порядка десятых долей миллирадиана. Трекер измеряет любую ошибку выравнивания между линией визирования на цель и вектором наведения оптической системы и выдает сигналы ошибки, которые дают команду сервоприводам системы скорректировать вектор наведения системы для достижения желаемого результата.
Для действительно эффективной системы управления огнем лазерный луч, направленный на удаленную цель, должен быть направлен на телевизионную систему и/или систему слежения FLIR. Предыдущие системы прицеливания включают те, которые наводят лазерный целеуказатель при отделении от пусковой установки (наземного, морского или воздушного базирования), например, только во время первоначальной сборки или через запланированные интервалы времени в ремонтной мастерской. Другие системы позволяют вести прицеливание, когда лазерный блок установлен на ракете-носителе. Однако эти системы предшествующего уровня техники ограничены случайным прицеливанием на безопасном для лазера расстоянии или прицеливанием по траектории полета для каждой миссии летательного аппарата. Система, которая допускает наименьшую ошибку прицеливания во многих миссиях, — это система, основанная на бортовом прицеливании. Методы бортового прицеливания могут включать выравнивание оптической оси лазера только в начале миссии в ответ на команду, инициированную пилотом, или прицеливание может инициироваться каждый раз, когда активируется система управления огнем.
Примеры известных методов прицеливания приведены в патенте США No. № 3628868, выданный 21 декабря 1971 г. на имя Starkey и U.S. № 3752587, выданный 14 августа 1973 г. на имя Meyers et al. Старки показывает лазерное устройство прицеливания, которое имеет телескоп, установленный на корпусе лазера, и выполняет прицеливание с помощью ручной микрометрической регулировки. Мейерс описывает устройство прицеливания, в котором используется полоска материала, на которую направляется лазер во время операции прицеливания. Лазер прожигает отверстие в полосе, позволяя свету проходить к телевизионному датчику. Созданное таким образом изображение выравнивается на телевизионной камере посредством ручной регулировки горизонтального и вертикального потенциометров, которые центрируют изображение относительно оптического перекрестия. Ни в одной из этих ссылок не раскрывается автоматическая ориентация, и этот факт имеет большое значение, когда осознается, что пилот, например, занят полетными задачами самолета и в таких условиях не может точно и надежно выполнить ручную лазерную ориентацию.
Патент США. № 4 155 096, выданный 15 мая 1979 г. Томасу и др., Обучал автоматическому лазерному прицеливанию. Эти патентообладатели добились наведения лазера системы лазерного целеуказания на нулевую точку автоматического телевизионного трекера, выборочно вызывая обратное отражение лазерного луча на видеодатчик системы, который взаимодействует с телевизионным трекером. Трекер фиксируется на отраженном назад лазерном пятне, а сигналы ошибки трекера используются в контуре управления с обратной связью для управления смещением растра видеодатчика. Напряжения смещения растра центрируют развертки видео вокруг пятна лазера, тем самым обнуляя сигналы ошибки трекера и автоматически обеспечивая прицеливание с помощью лазера.
Хорошо известно, что некоторые ракеты предназначены для запуска с наземного, водного или летательного аппарата и последующего наведения на выбранную цель с помощью оптического наведения, радиолокационного наведения и т.п. Одна такая система, представляющая интерес для настоящего изобретения, включает в себя наземное транспортное средство, имеющее несколько пусковых труб для ракет с реактивными двигателями, причем эти ракеты наводятся на цель с помощью системы наведения по лучу.
В средствах слежения за ракетой «земля-воздух» могут, например, использоваться телевизионные датчики, а также датчики FLIR (инфракрасные датчики переднего обзора), установленные на ракете-носителе, чтобы обеспечить возможность отслеживания цели, например самолета, в дневное время, как а также в условиях плохой видимости. На таком транспортном средстве находятся не только эти компоненты, но и множество зум-оптических систем, так что ракета может точно отслеживаться первой оптической подсистемой, а затем концентрированная информация наведения отправляется на ракету второй оптической подсистемой во время полета ракеты. фаза горения мотора, когда шлейф от мотора трудно пробить. Затем третья оптическая подсистема обеспечивает конечное наведение во время фазы без двигателя или на этапе движения ракеты по побережью, когда точные команды наведения на ракету чрезвычайно важны для перехвата цели.
Как подробно объяснялось в упомянутом выше изобретении Амона и Массона, устройство Zoom Projection Optic (ZPO) представляет собой систему наведения луча электромагнитного излучения, которая пространственно кодирует поперечное сечение луча наведения для создания большого количества элементов разрешения Каждый элемент разрешения однозначно обозначается цифровым кодом, осуществляемым частотной модуляцией излучения в каждом элементе разрешения в соответствии с другим цифровым словом. Другими словами, создается «коридор наведения», позволяющий ракете непрерывно получать сигналы вверх/вниз и влево/вправо и осуществлять коррекцию траектории полета ракеты по центральному разрешающему элементу матрицы элементов. Для конечного наведения ракеты используется оптический прибор ЗПО, через который направляется лазерная энергия.
Другими словами, создается «коридор наведения», позволяющий ракете непрерывно получать сигналы вверх/вниз и влево/вправо и осуществлять коррекцию траектории полета ракеты по центральному разрешающему элементу матрицы элементов. Для конечного наведения ракеты используется оптический прибор ЗПО, через который направляется лазерная энергия.
Устройство ZPO предпочтительно используется в сочетании с парой колесиков сетки, вращающихся в противоположных направлениях, которые используются для пространственного кодирования поперечного сечения луча наведения для создания большого количества элементов разрешения, используемых при окончательном наведении ракеты. Более подробную информацию о таких колесах сетки можно найти в патенте США на имя Аллена С. Лейтона, патент США. № 4 299 360 от 10 ноября 1981 г. Во время прицеливания эти колеса прицеливания располагаются в заранее установленном стационарном положении для определения линии прицеливания с высокой точностью. Этот оптический путь используется для выравнивания других оптических компонентов системы, чтобы обеспечить правильное направление визирования.
Настоящее изобретение было разработано в результате усилий по быстрому и высокоточному достижению направления визирования.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В соответствии с этим изобретением мы разработали устройство прицеливания, легко адаптируемое для встраивания в башню такого типа, который можно легко перевозить на транспортном средстве, таком как наземное или водное транспортное средство. транспортное средство на базе. Такая турель включает в себя первое средство крепления для поддержки множества световозвращающих оптических блоков в близко расположенном ряду и второе средство крепления, которое может вращаться по высоте, а также поворачиваться по азимуту. Во втором монтажном средстве или вращательном оптическом узле установлены зум-проекционная оптика (ZPO), ТВ-трекер, переднее инфракрасное устройство (FLIR) и командная оптика. Командная оптика включает зум ракетного слежения (MTZ) и лазерную оптику временного режима (TMLO), а также лазер, используемый в сочетании с такими компонентами. Командная оптика предназначена для отслеживания и наведения ракеты с лучевым наведением в период работы ракетного двигателя ракеты, когда использование ЗПО может быть не столь эффективным.
Командная оптика предназначена для отслеживания и наведения ракеты с лучевым наведением в период работы ракетного двигателя ракеты, когда использование ЗПО может быть не столь эффективным.
Вращающийся оптический узел или второе средство крепления обычно имеют цилиндрическую форму, при этом главная ось цилиндра обычно расположена горизонтально. Из-за своей конфигурации мы часто называем вращающуюся оптическую сборку «пепельницей». Именно вокруг такой горизонтальной оси пепельница или второе средство крепления могут вращаться для компенсации изменения высоты, при этом весь оптический узел может вращаться вокруг вертикальной оси через пьедестал, установленный на башне транспортного средства, когда требуется переместить оптические компоненты. по азимуту.
Основной оптической осью для прицеливания является ось ZPO, по которой информация об азимуте и угле места, полученная с помощью лазера, взаимодействующего с вращающимися в противоположных направлениях колесами прицельной сетки, передается ракете, наводимой на цель. Иными словами, таким образом определяется первичный коридор наведения, по которому наводится наводящаяся ракета до столкновения с целью. Мы рассматриваем лазер, работающий совместно с ЗПО, и скрещенные щели прицельных колес (создаваемые при расположении колес в заранее установленное стационарное положение) как определяющее интегрированную лазерную систему. Мы могли бы, конечно, модулировать выход лазера, чтобы обеспечить наведение ракеты на всех этапах полета, но на этапе полета после выгорания двигателя, когда используется ZPO, мы предпочитаем использовать вращающиеся колеса сетки, потому что такое использование позволяет получать очень точную информацию о местоположении.
Иными словами, таким образом определяется первичный коридор наведения, по которому наводится наводящаяся ракета до столкновения с целью. Мы рассматриваем лазер, работающий совместно с ЗПО, и скрещенные щели прицельных колес (создаваемые при расположении колес в заранее установленное стационарное положение) как определяющее интегрированную лазерную систему. Мы могли бы, конечно, модулировать выход лазера, чтобы обеспечить наведение ракеты на всех этапах полета, но на этапе полета после выгорания двигателя, когда используется ZPO, мы предпочитаем использовать вращающиеся колеса сетки, потому что такое использование позволяет получать очень точную информацию о местоположении.
Как правило, соответствующие выходные окна FLIR, телевизора и других компонентов располагаются примерно на одинаковом расстоянии от осей вращения, так что при перемещении вращающегося узла по высоте или азимуту несколько окон перемещаются на одинаковые величины. Из-за необходимости наведения ТВ, FLIR и командной оптики (включая МТЗ и ТМЛО) на ЗПО, например, ежедневно, желательно использовать фиксированные оптические компоненты, известные как ретрорефлекторные сборки или призменные сборки, которые должны быть легко доступны для прицеливания по мере необходимости. Вместо того, чтобы на вспомогательном транспортном средстве перевозить юстировочные узлы или призмы или устанавливать их каждый раз, когда необходимо прицеливание, мы вместо этого выделяем часть турели, примыкающую к поворотному оптическому узлу, для установки ретрорефлекторных узлов. Эти узлы установлены на том, что мы называем оптическим основанием модуля прицеливания, также известным как первое средство крепления. Затем, когда необходимо прицеливание, необходимо только повернуть пепельницу или второе средство крепления вокруг своей горизонтальной оси, вверх и вокруг в обычное заднее положение так, чтобы оно было обращено к части башни, содержащей узлы прицеливания, таким образом позволяя операция быстрого прицеливания.
Вместо того, чтобы на вспомогательном транспортном средстве перевозить юстировочные узлы или призмы или устанавливать их каждый раз, когда необходимо прицеливание, мы вместо этого выделяем часть турели, примыкающую к поворотному оптическому узлу, для установки ретрорефлекторных узлов. Эти узлы установлены на том, что мы называем оптическим основанием модуля прицеливания, также известным как первое средство крепления. Затем, когда необходимо прицеливание, необходимо только повернуть пепельницу или второе средство крепления вокруг своей горизонтальной оси, вверх и вокруг в обычное заднее положение так, чтобы оно было обращено к части башни, содержащей узлы прицеливания, таким образом позволяя операция быстрого прицеливания.
В интересах создания сборок ретрорефлекторов по разумной цене мы используем отдельные узлы визирования, расположенные на близком расстоянии друг от друга. После поворота в направленное назад положение по оси визирования пепельницу можно поворачивать последовательно по мере необходимости при наведении по трем оптическим путям. Мусорный бак поворачивается в несколько разные положения для достижения совмещения с первым и вторым узлами ретрорефлектора, а затем поворачивается в еще одно другое положение для достижения совмещения с третьим узлом ретрорефлектора. Эти последовательные этапы прицеливания выполняются быстро, но с высокой точностью.
Мусорный бак поворачивается в несколько разные положения для достижения совмещения с первым и вторым узлами ретрорефлектора, а затем поворачивается в еще одно другое положение для достижения совмещения с третьим узлом ретрорефлектора. Эти последовательные этапы прицеливания выполняются быстро, но с высокой точностью.
Основной задачей нашего изобретения является обеспечение возможности прицеливания для любой системы слежения за ракетами с радиусом действия от 3 мкм до 5 мкм, в которой в основном используется лазер CO 2 .
Еще одной важной целью нашего изобретения является создание нового узла оптических устройств наведения, примыкающего к поворотному оптическому узлу, содержащему множество оптических средств слежения, так чтобы можно было легко и удобно выполнять наведение устройств слежения на лазерную систему наведения.
Еще одной важной целью настоящего изобретения является создание в нашем новом узле направления направления средства, с помощью которого можно осуществлять преобразование длины волны, чтобы обеспечить быстрое направление направления оптических датчиков, работающих в различных частях оптического спектра.
Еще одна цель нашего изобретения состоит в том, чтобы сделать доступным устройство, использующее вращающиеся колеса прицеливания, обеспечивающие наведение ракеты, устройство прицеливания при использовании в сочетании с такими колесами прицеливания, расположенными в заранее установленных стационарных положениях, так что различные оптические датчики может быть направлен на очень точную линию визирования.
Еще одной целью нашего изобретения является создание нового способа наведения множества оптических путей, используемых в системе наведения ракетного луча, к используемым в ней устройствам слежения за целями.
Эти и другие цели, признаки и преимущества настоящего изобретения станут более очевидными по мере дальнейшего описания.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
РИС. 1 представляет собой вид в перспективе типичного поворотного оптического узла или мусорного бака, используемого на башне транспортного средства, в котором содержатся компоненты, используемые для наведения ракет класса «земля-воздух» или «земля-земля» на их соответствующие цели;
РИС. 2 представляет собой вид сбоку оптического основания модуля прицеливания, на котором расположены несколько ретрорефлекторов, используемых для прицеливания нескольких систем наведения и слежения за ракетами, содержащихся в части башни, показанной на фиг. 1;
2 представляет собой вид сбоку оптического основания модуля прицеливания, на котором расположены несколько ретрорефлекторов, используемых для прицеливания нескольких систем наведения и слежения за ракетами, содержащихся в части башни, показанной на фиг. 1;
РИС. 3 представляет собой вид сбоку задней стороны оптической платформы по фиг. 2, на которой конструкция и расположение трех сборок ретрорефлекторов, используемых в соответствии с настоящим изобретением, показаны с некоторыми подробностями;
РИС. 4 представляет собой вид в перспективе, на котором вращающийся оптический узел или мусорный бак повернут примерно на 180° от положения, показанного на фиг. 1, с мусорным баком в этом новом положении, показанном в типичном взаимодействии направления визирования с одним из ретрорефлекторных узлов, которым является ретрорефлекторный узел ZPO-FLIR;
РИС. 5a-5c представляют собой схематический вид сверху поворотной оптической сборки, последовательно взаимодействующей с узлами ретрорефлекторов ZPO-TV, ZPO-FLIR и ZPO-Command Optic;
РИС. 6а представляет собой вид сбоку пары колесиков сетки, используемых в фокальной плоскости проекционной оптики с масштабированием;
6а представляет собой вид сбоку пары колесиков сетки, используемых в фокальной плоскости проекционной оптики с масштабированием;
РИС. 6b представляет собой вид сбоку, аналогичный фиг. 6а, на котором колеса сетки повернуты в положение визирования; и
ФИГ. 7a-7c представляют собой несколько идеализированные виды узлов ретрорефлекторов, позволяющих наводить устройства TV, FLIR и Command Optics на ZPO, при этом в некоторых из этих узлов используются важные преобразования длины волны.
ПОДРОБНОЕ ОПИСАНИЕ
Обращаясь сначала к РИС. 1 видно, что мы изобразили часть башни транспортного средства, оснащенную множеством труб 10 для пуска ракет, таких как ракеты класса «земля-воздух» или ракеты класса «земля-земля». Между двумя рядами трубок установлен вращающийся оптический узел 12, в котором используется основная часть этого изобретения.
Вращающийся оптический узел 12 обычно имеет цилиндрическую форму и расположен своей главной осью, как правило, в горизонтальной плоскости, и из-за его внешнего вида его часто называют «пепельницей». Мусорный бак может вращаться вокруг своей горизонтальной оси, так что он может легко изменять угол подъема, а также может поворачиваться вокруг своей подставки 16. Радарная антенна 18 также может быть установлена на башне транспортного средства, но это не имеет прямого отношения к настоящему изобретению.
Мусорный бак может вращаться вокруг своей горизонтальной оси, так что он может легко изменять угол подъема, а также может поворачиваться вокруг своей подставки 16. Радарная антенна 18 также может быть установлена на башне транспортного средства, но это не имеет прямого отношения к настоящему изобретению.
На передней части мусорного бака 12 расположено множество окон или проемов. Первое из них мы называем окном ZPO 20, поскольку оно относится к оптическому проецированию с увеличением, используемому для формирования основного оптического пути, по которому наводится каждая ракета. Также изображено окно 22, используемое совместно с телевизором, которое легко может распознавать контраст цели по отношению к фону. Кроме того, мы используем окно 24 FLIR, последнее относится к устройству «переднего обзора в инфракрасном диапазоне», используемому в башне для отслеживания цели, такой как самолет, танк или другая горячая цель. Дополнительно используется окно командной оптики 26. Как упоминалось ранее и как будет объяснено более подробно в дальнейшем, мы используем термин «командная оптика» для обозначения нашей новой лазерной оптики временного режима (TMLO) и нашей новой системы слежения за ракетами (MTZ). ) устройства, объединенные по существу в один корпус.
) устройства, объединенные по существу в один корпус.
Устройства TMLO, MTZ и ZPO подробно описаны в одновременно находящейся на рассмотрении патентной заявке Макса Амона и Андре Массона, цитируемой выше, и, поскольку она представляет собой подробное описание командной оптики, в данном случае она считается ненужной. подробно описать эти компоненты. Следует четко понимать, что все соответствующие идеи этой патентной заявки включены в настоящую заявку посредством ссылки.
Значительное количество мешающего инфракрасного излучения генерируется двигателем ракеты во время запуска, поэтому мы обычно резервируем использование оптики ZPO для конечного наведения и используем командную оптику для отправки и получения информации о местоположении в начальный период. полета ракеты, в то время как реактивный двигатель ракеты продолжает гореть, поскольку в это время необходим концентрированный луч для проникновения в шлейф двигателя. Компонент Missile Tracker Zoom (MTZ) командной оптики служит для постоянного отслеживания положения ракеты во время полета с двигателем, в то время как TMLO предоставляет информацию о местоположении ракеты во время периода работы двигателя, поскольку он обеспечивает очень концентрированный луч, который способен проникать в шлейф двигателя.
Как должно быть очевидно, очень важно, чтобы различные компоненты и устройства мусорной корзины — FLIR, телевизор, командная оптика и зум-проекционная оптика — были ориентированы по оси, чтобы эти компоненты могли эффективно и точно взаимодействовать и сотрудничать вместе. С этой целью в соответствии с настоящим изобретением мы разработали устройство, позволяющее легко и точно наводить на эти устройства и компоненты без необходимости установки какого-либо дополнительного оборудования.
На транспортном средстве, как правило, за вращающимся оптическим узлом или мусорным баком установлена опорная панель 27, угол которой показан на РИС. 1. Панель 27 служит опорой для некоторых электронных систем, а также поддерживает оптическую платформу 28 модуля прицеливания, передняя и задняя стороны которой подробно показаны на фиг. 2 и 3 соответственно. Оптическая платформа визирования использует несколько ретрорефлекторов, применяемых в соответствии с настоящим изобретением, и здесь она также упоминается как первое монтажное средство. Внешний вид оптической платформы 28, если смотреть из мусорного бака 12, если смотреть назад, показан на фиг. 2, тогда как на фиг. 3, где показана задняя сторона оптического основания 28 модуля прицеливания, показаны многие из фактических компонентов отдельных узлов ретрорефлекторов. В этом контексте мы называем пепельницу или поворотный оптический узел вторым средством крепления. Хотя мы не должны ограничиваться какой-либо одной конструктивной компоновкой, поскольку речь идет о компонентах сборок ретрорефлекторов, мы предпочитаем использовать трубки из инвара, обычно диаметром два дюйма, в которых монтируются конкретные оптические компоненты, составляющие каждый из световозвращатели.
Внешний вид оптической платформы 28, если смотреть из мусорного бака 12, если смотреть назад, показан на фиг. 2, тогда как на фиг. 3, где показана задняя сторона оптического основания 28 модуля прицеливания, показаны многие из фактических компонентов отдельных узлов ретрорефлекторов. В этом контексте мы называем пепельницу или поворотный оптический узел вторым средством крепления. Хотя мы не должны ограничиваться какой-либо одной конструктивной компоновкой, поскольку речь идет о компонентах сборок ретрорефлекторов, мы предпочитаем использовать трубки из инвара, обычно диаметром два дюйма, в которых монтируются конкретные оптические компоненты, составляющие каждый из световозвращатели.
Мы рассматриваем зум-проекционную оптику, включая колеса сетки, используемые в фокальной плоскости ZPO в сочетании с освещающим лазером, как определяющие основную линию визирования (LOS) на цель, поэтому несколько апертур узлов ретрорефлекторов каждый представлен в оптической связи с апертурами ZPO на фиг. 2, где они сгруппированы в центральной части оптической платформы 28 модуля прицеливания. Апертура 30а на фиг. 2 связан с ретрорефлектором 32 направления, используемым для наведения ТВ-трекера на ЗПО; апертура 30b связана со световозвращателем 34, используемым для наведения трекера FLIR на ZPO; а апертура 30с связана со световозвращателем 36, используемым для наведения командной оптики на ZPO.
2, где они сгруппированы в центральной части оптической платформы 28 модуля прицеливания. Апертура 30а на фиг. 2 связан с ретрорефлектором 32 направления, используемым для наведения ТВ-трекера на ЗПО; апертура 30b связана со световозвращателем 34, используемым для наведения трекера FLIR на ZPO; а апертура 30с связана со световозвращателем 36, используемым для наведения командной оптики на ZPO.
Лазер, используемый в пепельнице или во втором средстве крепления для обеспечения наведения ракет по лучу, может представлять собой лазер CO 2 , и этот лазер используется во время процедуры прицеливания для последовательного направления лазерной энергии в каждый из ретрорефлекторов прицеливания сборки. В частности, при прицеливании с помощью ретрорефлектора ЗПО-ТВ энергия лазера направляется в апертуру 30а; при наведении световозвращателя ZPO-FLIR такая энергия направляется в апертуру 30b; а при прицеливании с использованием ретрорефлектора Command Optics такая энергия направляется в апертуру 30с. Мы рассматриваем лазер, работающий совместно с ZPO, когда колеса сетки неподвижны со скрещенными прорезями, как определение интегрированной лазерной системы. Позиционирование колёс визирной сетки во время процедуры визирования будет обсуждаться в связи с фиг. 6а и 6б.
Мы рассматриваем лазер, работающий совместно с ZPO, когда колеса сетки неподвижны со скрещенными прорезями, как определение интегрированной лазерной системы. Позиционирование колёс визирной сетки во время процедуры визирования будет обсуждаться в связи с фиг. 6а и 6б.
На фиг. 3 мы проиллюстрировали внешний вид узлов ретрорефлектора оси прицеливания, и на этом рисунке видны некоторые важные компоненты. Видны корпус 38 параболического зеркала 68, связанного с узлом 32 телевизионного ретрорефлектора, а также электрический провод 40, связанный с лампой накаливания или лампочкой (не показана), установленной в параболическом отражателе, причем эта лампочка используется для причина, которая будет обсуждаться в дальнейшем. Также видно на фиг. 3 показан корпус 44 потолочного зеркала 48, используемого в сборке FLIR-ZPO 34, и корпус 46 параболического зеркала 82, используемого в сборке ZPO-Command Optics 36.
Когда пепельница должна работать в режиме прицеливания, она поворачивается вверх вокруг своей горизонтальной оси до тех пор, пока не станет направленной назад. ИНЖИР. 4 показан вращающийся оптический узел или мусорный бак в его направленном назад режиме прицеливания, где в данном случае он взаимодействует с узлом световозвращателя ZPO-FLIR 34. Как видно из этого рисунка, ближний конец этого световозвращателя использует так называемое зеркало крыши 48, внутренние поверхности которого находятся под углом 9Угол 0° и посеребрение. Отражатель на дальнем конце этого узла представляет собой плоское зеркало 74.
ИНЖИР. 4 показан вращающийся оптический узел или мусорный бак в его направленном назад режиме прицеливания, где в данном случае он взаимодействует с узлом световозвращателя ZPO-FLIR 34. Как видно из этого рисунка, ближний конец этого световозвращателя использует так называемое зеркало крыши 48, внутренние поверхности которого находятся под углом 9Угол 0° и посеребрение. Отражатель на дальнем конце этого узла представляет собой плоское зеркало 74.
Теперь обратимся к фиг. 5а-5с видно, что мы здесь схематически изобразили пепельницу или поворотную оптическую сборку, используемую в ее первом рабочем режиме, в котором она используется последовательно в положениях, где прицеливание ZPO-TV; прицеливание ZPO-FLIR; и прицеливания ZPO-Command Optics.
На фиг. 6a и 6b мы проиллюстрировали пару колесиков сетки 54 и 56 того типа, которые, как подробно пояснено в ранее упомянутой патентной заявке Амона и Массона, используются в фокальной плоскости Zoom Projection Optics. Эти колеса изготовлены из нержавеющей стали, чтобы они могли выдерживать значительный эффект нагрева, вызванный использованием лазера для освещения.
Как поясняется в патенте США Layton № 4 299 360, колеса сетки содержат определенную информацию, которая проецируется на ракету для передачи точной информации о местоположении. В частности, за счет размещения определенных кодированных прорезей на внешних частях колесиков сетки лазерный луч прерывается таким образом, чтобы обеспечить точную информацию о местоположении ракеты, наводимой на цель. Мы предпочитаем, чтобы рубленый луч создавал матрицу размером 16 на 16 ячеек, где каждая ячейка имеет, скажем, 3/4 метра на стороне. Таким образом, зум-проекционная оптика служит для создания матрицы ячеек постоянного размера 12 на 12 метров0 во время полета ракеты после выгорания двигателя, что достигается с помощью возможности масштабирования. Благодаря двум приемникам заднего обзора, используемым на ракете, наводимой на цель, система наведения ракеты способна декодировать спроецированную схему и, как следствие, заставить ракету двигаться к центральной ячейке матрицы. . Только когда ракета движется по центру проецируемого лазерного коридора, она не будет получать сигналы, требующие ее движения вверх или вниз, вправо или влево. Родственное изобретение Макса Амона и Клиффорда Люти под названием «Окно МДП», поданное 21 мая 1919 г.84.
Родственное изобретение Макса Амона и Клиффорда Люти под названием «Окно МДП», поданное 21 мая 1919 г.84.
Сер. № 612,194 касается значительных частей окон ствольной коробки ракеты.
Узел колеса энкодера в основном состоит из сегмента 50 колеса энкодера с вертикальным разрешением и сегмента 52 колеса энкодера по горизонтали; см. фиг. 6а и 6б. Каждое колесо кодирующего устройства 54 и 56 подходящим образом соединено с соответствующей ведущей шестерней (не показано). Вертикальная ведущая шестерня и горизонтальная ведущая шестерня находятся в зацеплении и приводятся в желаемое встречное вращение предпочтительно одним двигателем. С этой целью двигатель (не показан) приводит в зацепление одну из ведущих шестерен. Сегменты энкодера 50 и 52 занимают менее 180 градусов каждый. Таким образом, их можно заставить вращаться, предпочтительно по одному, в лазерном луче, при этом сегменты 50 и 52 не перекрываются в области луча. Вращение в этом случае может происходить в направлении стрелок, показанных на колесных элементах 54 и 56 на фиг. 6а.
6а.
Чтобы упростить первоначальную настройку лазера, мы обеспечиваем сравнительно большое, как правило, круглое отверстие 57 вблизи периферии каждого из колес сетки, как лучше всего видно на ФИГ. 6б. Затем, когда колеса находятся в состоянии покоя в положении, показанном на фиг. 6а, лазерный луч может легко беспрепятственно проходить через эти совмещенные круглые отверстия.
Несмотря на то, что диски вращаются в противоположных направлениях с постоянной скоростью во время передачи информации наведения на ракету, они должны оставаться неподвижными во время процедуры прицеливания. В каждом диске вырезана короткая окружная прорезь для прицеливания, это прорезь 58 в колесе 54 и прорезь 59.в колесе 56, как лучше всего видно на фиг. 6а. Таким образом, когда диски останавливаются в положении, показанном на фиг. 6b, можно легко выполнить прицеливание. Перекрещенные прорези (или прорези) в сочетании с ZPO образуют самое основное определение нашей линии прямой видимости (LOS) до цели.
Обратимся теперь к фиг. 7a, мы показали в несколько упрощенном виде, как телевизор наведен на зум-проекционную оптику. Важно отметить, что необходимо выполнить преобразование длины волны, чтобы телевизор мог видеть энергию лазера во время процедуры прицеливания. Для этого используется дихроичный светоделитель 63, в центре которого расположена мишень, покрытая жидкокристаллическим слоем 64. Таким образом, лазерная энергия, прошедшая через оптику ZPO, отражается зеркалом 66 и проходит через дихроичный 63, который прозрачен для энергии 10,6 мкм. Затем эта энергия попадает на параболический отражатель 68, который сконфигурирован и размещен таким образом, чтобы сфокусировать лазерную энергию на жидкокристаллической мишени 64. Тепло, выделяемое при поглощении, заставляет жидкий кристалл реагировать, образуя темное пятно. Благодаря колбе 70, которая направляет свет через линзу 72, расположенную в центре параболического рефлектора 68, обеспечивается яркий фон, позволяющий телевизионному датчику легко видеть темное пятно, образовавшееся на кристаллическом слое.
Термоэлектрический охладитель (не показан) используется для управления температурой мишени жидкого кристалла, что обеспечивает требуемую чувствительность жидкого кристалла.
После того, как телевизор обнаружит темное пятно, необходимо переместить видеорастр так, чтобы телевизионная сетка совпала с темным пятном жидкокристаллической мишени. Когда это сделано, ТВ выравнивается по линии прямой видимости ЗПО.
Несколько похожая операция теперь используется для выравнивания FLIR с оптикой Zoom Projection. Следует отметить, что в настоящее время преобразование длины волны не требуется, поскольку трекер FLIR чувствителен к энергии с длиной волны 10,6 мкм, излучаемой лазером.
Как показано на фиг. 7b, мы предпочитаем использовать ретрорефлектор, содержащий ранее упомянутое зеркало 48 крыши, посеребренные внутренние поверхности которого образуют между собой прямой угол. Энергия лазера, выходящая из оптики ZPO, сначала отражается верхним зеркалом, а затем отражается плоским зеркалом 74 в датчик FLIR. Трекер FLIR теперь отслеживает инфракрасное изображение поперечных щелей и наводит на FLIR путем электронного смещения растра.
Наконец, что касается ФИГ. 7а-7с, на фиг. 7с следует понимать, что нам необходимо добиться направления как МТЗ, так и ТМЛО. На этом рисунке мы схематически изобразили лазер 90, обычно лазер CO 2 , направляя свою энергию через скрещенные прорези колесиков 54 и 56. .) Эта энергия лазера проходит через оптику ZPO и сначала попадает на зеркало 76, которое служит для направления лазерной энергии через дихроичный светоделитель 78. Этот дихроичный светоделитель был выбран таким образом, чтобы примерно 50% энергии 10,6 мкм от лазер проходит сквозь него и отражается параболическим отражателем 82. Примерно в центре дихроичного светоделителя расположена тонкая полимерная пленка 80, покрытая сажевой краской. Мы предпочитаем каптоновый пластик, но не ограничиваемся им. Лазерная энергия, проходящая через дихроичный светоделитель 78, отражается параболическим зеркалом 82 и фокусируется на полимерной мишени 80. Каптоновый пластик поглощает лазерную энергию и излучает в диапазоне длин волн от 3,5 мкм до 4,2 мкм. Это излучение реколлимируется параболическим отражателем, затем отражается от дихроичного светоделителя и затем поступает через зеркало 39., оптика мтз. (Зеркало 39 находится вне плоскости бумаги и соответствует карданному зеркалу 9 из патентной заявки Амона и Массона, касающейся командной оптики.) Затем зеркало 39 настраивается таким образом, чтобы импульсы, создаваемые вращающимся оптическим клином МТЗ вызывает получение детектором МТЗ равномерно расположенных импульсов света.
Как следует из заявки на патент Амона и Массона, касающейся Command Optics, в тракте MTZ используется детектор 11. Благодаря вращающемуся клину на детектор проецируется продолговатое световое пятно, при этом это пятно или капля света движется по кругу вокруг четырех чувствительных стержней детектора. Каждый из этих стержней расположен радиально и расположен под углом 9интервалы 0°. Однако детектор и клин здесь не изображены.
При центральном расположении этого светового пятна будут приниматься четыре равноотстоящих друг от друга выходных импульса одинаковой высоты, а при смещении оси МТЗ от намеченной линии визирования к цели будут приниматься импульсы с переменным интервалом. Относительные промежутки между импульсами указывают направление смещения MTZ от линии визирования ZPO и диктуют изменение положения зеркала 39. MTZ можно рассматривать как направленный по направлению к ZPO, когда выходные импульсы равномерно распределены.
Для прицеливания оптики TMLO зеркало 86 (соответствующее зеркалу 3 заявки на патент Amon and Masson «Command Optics») переключается обратно так, что оно направляет энергию лазера на зеркало 88, что в свою очередь направляет эту энергию на дихроичный светоделитель 78. Примерно 50% этой энергии направляется на параболический отражатель 82, служащий для фокусировки лазерной энергии на полимерную мишень 80, которая, как и прежде, излучает в диапазоне от 3,5 мкм до 4,2 мкм. диапазон длин волн. Это излучение отражается параболическим зеркалом 82 и, как и прежде, попадает в детектор МТЗ.
Часто обнаруживается, что положение зеркала 39 для достижения направления визирования через оптику ZPO отличается от положения зеркала для достижения направления визирования TMLO. Другими словами, замкнутый сервоконтур зеркала на карданном подвесе может обеспечить два совершенно разных показания для наведения TMLO и ZPO на ось MTZ.
Хотя возможны и другие решения, наш предпочтительный вариант — отметить несоответствие между линиями визирования ТМЛО и МТЗ, а затем компенсировать это несоответствие в полете ракеты соответствующими входными данными в системное программное обеспечение.
Теперь должно быть очевидно, что мы предоставили очень выгодное устройство модуля прицеливания и метод, с помощью которого можно быстро и точно выполнить прицеливание оптической системы и наиболее удобным способом.
Трактор МТЗ-82: манилебау, дисгрифиад, прис
г. в. 1974, даэт и трактор МТЗ-82 одди ар линелл и цинуллиад, а даэт у нодведдион технегол охоно и йматэб тэйлвнг и’р гальв циниддоланна амейпудфил амэйтйаддолу, амаэтиаддолу адейладу. Er mwyn gwneud y трактор yn well na’i ragflaenwyr, roedd yn rhaid i’r craftyyr weithio’n galed. Y prif wahaniaeth или модель blaenorol oedd modur newydd sy’n caniatáu i’r трактор ddatblygu cyflymder da ar wahanol fathau o balmant a gweithio gydag amrywiaeth o atodiadau. Roedd Gwasgariad newydd y трактор yn rhoi golwg fwy modern a mwy iddi, ac roedd gan y cab modern fod yn waith cynhyrchiol. Heddiw, byddwn yn adolygu prif nodweddion technegol y трактор МТЗ-82 («Беларвс») a darganfod beth sy’n penderfynu ei boblogrwydd.
Gwybodaeth gyffredinol
Mae’r трактор MTZ-82, y mae ei nodweddion technegol y byddwn yn dadansoddi isod, mewn saw ffordd yn union yr unfath â’i ragflaenydd, MTZ-80. Yn wahanol i’r model hen, mae ganddi yrru pedair olwyn ac awydd eithaf cymedrol.
Yn allanol ac yn dechnegol, maer model yn cyd-fynd yn llawn â’i deulu, y prif nodweddion nodedig yw adeiladu lled-ffrâm, gwahanol ddiamedr yr olwynion (y cefn yn llawer mwy blaen), a’r modur sydd wedi’i leoli o flaen y caban.
Y llinell
Yn ystod y broses o gynhyrchu трактор МТЗ-82, roedd nodweddion technegol wedi’u moderneiddion raddol, ymddangosodd nifer o fersiynau gwahanol, ac mae do do pob un ohonynt yn perfformio rhai swy. Er gwaethaf y ffaith bod yr addasiadau yn wahanol i’w gilydd ar gyfer nifer o nodweddion, maen nhw i gyd wedi’u cynhyrchu o dan yr enw MTZ-82 gydag ychwanegu mynegeion ychwanegol. Год выпуска 2000 г., майор трактора wedi’i gynhyrchu dan yr enw «Беларус-82».
Yn ychwanegol at y fersiwn sylfaenol, mae’r amrediad model yn cynnwys addasiadau или fath:
- МТЗ-82.1. Mae’r nodweddion technegol yr unfath, ond mae gan y caban gyfaint fwy.
- МТЗ-82.1-23/12. Fe’i gwahaniaethir gan olwynion blaen wedi’u hehangu.
- МТЗ-82Р. Wedi’i gynllunio ar gyfer cynaeafu reis.
- МТЗ-82Н. Wedi’i addasu i weithio ar y llethrau (mae’r sedd yn cymryd sefyllfa gyfforddus wrth newid y tir).
- МТЗ-82К. Wedi’i gynllunio i weithio ar lethrau serth. Wedi addasiad awtomatig o sefyllfa’r sgerbwd. Er mwyn sefydlogi’r peiriant, maegan y system gyswllt silindrau hydrolig ychwanegol.
- МТЗ-82Т. Wedi’i addasu ar gyfer casglu llysiau a gofal amdanynt. Oherwydd gerau olwyn mae ganddi glirio tir uwch.
Poblogrwydd
Mae presenoldeb nifer o addasiadau o’r fath yn nodi màs y трактор МТЗ-82. Nid nodweddion technegol y peiriant yw ei unig gerdyn trwm. Dadl bwyswl o blaid y трактор oedd ei brisel. Diolch i’r system gyrru all-olwyn, mae’r truck yr un mor llwyddiannus wrth ddelio ag amodau daearyddol a thyydd anodd. Gall yr addasiadau uchod weithio gyda 230 math o atodiadau ychwanegol. Mae’r rhain yn стоогометатели, копнозы, peiriannau torri, предлагают предлагать arbennig ac yn y blaen.
Peiriant a throsglwyddo
Yn dibynnu ar y flwyddyn gynhyrchu, roedd nodweddion technegol Трактор МТЗ-82 («Беларусь») yn amrywio. Hyd at 1985, roedd gan y car drosglwyddiad mecanyddol, a oedd yn cymryd 18/4 o gerau ar gyfer y gerau blaen a 16/4 ar gyfer yr olwynion cefn. Felly, roedd yn bosibl defnyddio’r pŵer modur yn effeithlon ar unrhyw gyflymder. Diolch i’r blychau предлагает гидро-реоли, roedd gan y gyrrwr yr opsiwn o newid gêr heb ddiffodd y cydiwr. Mae’r swyddogaeth hon ar gael ar gyfer modelau o ddiwedd y blynyddoedd cynhyrchu.
Mae gosodiadau hydrolig y cefn echel yn addasiadau newydd, y gellir eu cynnal o’r Pedal ac yn avtomatig. Mae патент ychwanegol y трактор yn ei gwneud yn bosibl cloi’r gwahaniaethol.
Mae injan дизель gyda chapas o 80 лошадиных сил yn caniatáu i’r трактор gyrraedd cyflymder o 35 км/ч. Yn ogystal, mae’n caniatáu ichi gydgrynhoi’r peiriant gyda phob math o atodiadau a gweithio’n ddi-dor gyda’r angen lleiaf ar gyfer cynnal a chadw. Mae’r peiriannau D-240 a D-243 yn cael eu cynhyrchu y Минский моторный завод. Mae’r injan pedwar-strôc yn cael ei yrru gan ddechreuwr trydan sydd â chyn-wresogydd. Mae’r modur yn cychwyn yr un mor dda yn ystod gwres yr haf a phryfed difrifol Chwefror.
Маеган и трактор размерами крино шир: 3810/2450/1970 мм.
Hydrolig
Mae’r трактор МТЗ-82, y mae ei nodweddion technegol yrydym yn eu trafod heddiw, â system hydrolig ar y cyfan. Mae’n cynnwys pwmp gêr, atgyfnerthiad hydrolig ar gyfer atodiadau, rheoleiddwyr safle a phŵer, dosbarthwr, silindr hydrolig sy’n rheoli’r cylchdro. Mae hyn i gyd yn cael ei weithredu o’r caban trwy gyfrwng liferi a phedlau.
Ходовая часть
Mae’r trefniant olwyn 2 × 2 yn sicrhau gweithrediad symly трактор. Yn yr olwynion blaen mae’r ataliad yn rhy anhyblyg, ac yn y cefn — ангиблыг. Mae’r rheswm dros hyn yn syml — mae’r olwynion blaen yn gyfrifol nid yn unig ar gyfer trosglwyddo Tork, ond hefyd am dro. Yn ystod gweithrediadau cludiant, gall yr echel flaen gael ei diffodd.
Gall y pellter rhwng olwynion y rhes gefn amrywio o 1,4 и 2,1 м. Gyda pellter cynyddol, mae sefydlogrwydd a phatentrwydd y cerbyd yn cynyddu. Maer trac blaen hefyd yn amrywio rhwng 1,2 и 1,8 м. Gall y cliriad amaethyddol fod yn wahanol, yn dibynnu ar fersiwn y трактор. Yn y fersiwn sylfaenol, mae’n 46,5 см.
System niwmatig
Mae’n cynnwys cywasgydd a dosbarthwr falf. Mae’r olaf yn cyflawni’r swyddogaeth o reoli breciau gwahaniaethol. Gellir defnyddio niwmateg y трактор ar gyfer pwmpio teiars, at ddibenion erill. Oherwydd nad yw’r system hon mor ddatblygedig ac yn gymhleth ag ychwanegion трамвай и трактор, mae’n llawer haws ei reoli.
Caban
O ystyried nodweddion technegol cyffredinol y трактор МТЗ-82, mae’n werth son am y caban. Mae’n werth nodi nad yw’n gysylltiedig â sgerbwd y трактор yn uniongyrchol. Caiff y caban ei osod ar siocledwyr rwber. Mae hyn yn lleihau dirgryniad a sŵn. Am yr un pwrpas, mae drysau’r car yn cael eu gorchuddio â haen o ddeunydd di-dor. Yn y caban, gallwch gynnal yr amodau tymheredd angenrheidiol oherwydd y posibilrwydd o wresogi ac awyru. Mae gwresogi yn digwydd oherwydd y gwres a gymerir o’r system hydrolig, ac awyru — trwy agor ffenestri a tho.
Cwmpas y cais
Gellir defnyddio’r peiriant hwn fel fforch godi, gwydr, cyfuno gwisgoedd, cyfrwng cludo a chloddio. Mae MTZ-82, y nodweddion technegol yr ydym yn eu trafod heddiw, yn beirant cyffredinol. Mae amlgyfundeb, ynghyd â chost isel, yn esbonio ei boblogrwydd. Gall ymhell или bob аналоговый dramor fod ag amrywiaeth mor eang o offer ychwanegol a chyda’r un llwyddiant i ymdopi â gwahanol dasgau. Хыд ын хын, маэ трактор МТЗ-82, й нодведдион технэгол ыр ыдым ын еу хадолыгу уход, мьюн цифлвр да туа 20 мил о ддолери. Gellir prynu model hynod, ond eithaf dibynadwy a hanner y pris.
МТЗ Беларусь | Страна 1262
Аки.в
Члан
- #25.221
Люди jel zna neko kako se puni sistem za hladjenje na 82, novi tip ima grejac u kabini? . .uglavnom kaplje mi antifriz na slavinici gde se otvara/zatvara grejanje .pa moram da ispustim antifriz da bi je zamenio ..e sad ne znam kako da napunim sistem ,tacnije nije mi jasno kako da napunim greiac kada je iznad hladnjaka
Jel ima ko skustva sa tim ? (термостат Мислим да немам)
Аттила
Члан
- № 25.222
Pa nikako to će sama da se napuni kad sastaviš sve napuniš hladnjak sa rashladnom technosću i upališ traktor i pustiš grejanje pa ce sama da se izluftira. Само после требования проверить технологичность у хладняку.
Аки.в
Члан
- #25. 223
Аттила je napisao(la):
Pa nikako to će sama da se napuni kad sastaviš sve napuniš hladnjak sa rashladnom technosću i upališ traktor i pustiš grejanje pa ce sama da se izluftira. Само после требования проверить технологичность у хладняку.
Кликните по проширению…
Nikako mi ne ide u glavu da ce se tecnost sama od sebe popeti gore u grejac, ali moram probati
Бобо
Члан
- #25.224
Pa kad upališ motor traktora vodna pumpa će tekućinu pumpati kroz sistem za grijanje… Kasnije samo u hladnjak nadotočiš koliko je sistem grijanja «povukao» i to je to.
МДрашко
Члан
- № 25.225
Aki.w je napisao(la):
Nikako mi ne ide u glavu da ce se tecnost sama od sebe popeti gore u grejac, ali moram probati
Нажмите на ссылку
Стави джой лотру и небрини.
Аки.в
Члан
- № 25. 226
Бобо je napisao(la):
Pa kad upališ motor traktora vodna pumpa će tekućinu pumpati kroz sistem za grijanje… Kasnije samo u hladnjak nadotočiš koliko je sistem grijanja «povukao» i to je to.
Кликните по проширению…
tako mi je i jedan prijatelj isto rekao .a jel smem da otvorim hladnjak kada mi je veksna za grejanje otvorena ,po nekoj logici sva tecnost koja je iznad hldnjaka ce iscureti .
jel njemu ulaz vode u grejac kabine sa desne strane kod pumpe ? или са леве код глава мотора?
Ристаа
Члан
- № 25.227
ne bi trebala isucreti voda kad je otvoren cep, vodu dobija sa termostata (или odakle vec na rusu ide) a vraca vodu u vodenu pumpu
Аттила
Члан
- № 25. 228
Sa leve strane ulazi voda ne treba da otvaraš čep kad radi traktor nego napuniš hladnjak pa zatvoriš i onda upališ traktor i pustiš ga da radi da bi počeo da greje u kabini posle kad ugasiš sačekaš malo da se hladi i dosipaš još tečnosti kolko fali . I posle još neki put Proriš nivo u hladnjaku kad je hladan motor.
вазовак
Члан
- № 25.229
Nebito je kuda dolazi a kuda se vraća .system spojenih sudova,kad bolje razmislim ni meni nije jasno kako voda iz hladnjaka u kabini ne iscuri kada se otvori hladnjak ,šta je sprečava kada je večinio0003
сремак84
Члан
- #25. 230
Bitno je,ne ide iz hladnjaka,vec sa vodene pumpe,s other strane termostata,kad he hladan termostat je zatvoren jel tako,zato i nije spojen sud,e kad je zagrejan nisamprovavao da li bi onda iz hladnjaka u kabini voda dizala nivo na cepu u hladnjaku…
Или сам я несто криво шватио…
ворчание
Члан
- #25.231
Dali je neko pravio pravio priključak na kompresoru Mtz-a za izduvavanje kabine или duvanje guma(brze spojke)?
френки
Члан
- № 25. 232
Ja imam ugradjeno dole na sudu gde je bio ventil za ispustanje kondenzovane vode.
Бранкович
Члан
- #25.233
Имам и я. У кабины сам сместио спойнику. Najviše zbog izduvavanja kabine. To sam uradio u oba traktora i bas je korisno.
Николлаа
Члан
- № 25. 234
Мозе нека слика како сте настили то, и мене занима патент?
вазовак
Члан
- № 25.235
сваки термостат има на себи едну малу рупику кой се не затвара,зато ми ние ясно како вода из кабине не излази
sremac84 je napisao(la):
Bitno je,ne ide iz hladnjaka,vec sa vodene pumpe,s other strane termostata,kad he hladan termostat je zatvoren jel tako,zato i nije spojen sud,e kad je zagrejan nisam proofravao da li bi onda iz hladnjaka u kabini voda dizala ниво на цепу у хладняку…
Или сам я несто криво шватио…Кликните по проширению.
декс
Члан
- #25.236
Kolega Brankovic je sigurno preneo patch iz kamiona
Za prasinu ne postoji bolje resenje!
Послато на SM-J710F с помошью Тапатока
Бранкович
Члан
- № 25. 237
@deks
Управо тако Прво сам видео у камину,а после и у тракторима
Всего:
Ичбинмилан
Члан
- № 25.238
nikollaa je napisao(la):
Moze neka slika kako ste namestili to, i mene zanima патент?
Кликните по проширению…
Ичбинмилан
Члан
- № 25. 239
Može li neko od kolega da mi da par informacija.
Интересуюсь изменением размера рупы на заднюю точку (центральную рупию) код МТЗ 50/52. Има 8 шрафова/рупа. Koliki je prečnik Centralnog otvora i koliko je rastojanje od centra jednog do centra otherog otvora za šraf ali popreko.
Da li pašu točkovi od 82?
савопунк
Запрещено
- #25.240
Pašu isto samo manja felna za 13.6 gumu.
МТЗ-82.1: характеристики и отзывы
Белорусское качество — тема особого разговора. Не секрет, что и по сей день изделия, выпускаемые в этой бывшей советской республике, славятся высочайшим качеством исполнения. И речь идет не только о продуктах питания и текстиле, но и о сельскохозяйственной технике. В этой статье мы подробно рассмотрим трактор МТЗ-82.1, его особенности, характеристики и отзывы пользователей.
История истории
Тракторостроение в Беларуси вышло на новый уровень в 1970-е годы, когда по постановлению Совета Министров СССР началось производство тракторов серии МТЗ. Именно в то время правительством было принято решение о выпуске очень мощного пропашного трактора. За основу был взят МТЗ-50. Конструкция машины претерпела многочисленные изменения, кабина также подверглась модернизации, обшивке, установлен новейший двигатель повышенной мощности. Первые испытания МТЗ-82.1 прошли в 1972 и оказался вполне удачным. На основе этих тестовых прогонов был сформирован перечень агрегированных узлов и деталей, а также создано около 230 устройств, которые были направлены на выполнение различных видов работ. При этом скорость трактора достигала 30 км/ч, что позволяло использовать его для перевозки.
Первая ласточка
Первая описываемая машина была выпущена Минским заводом в 1974 году. Уже на начальном этапе эксплуатации пользователи стали очень положительно отзываться о тракторе, и инженеры решили и дальше увеличивать объемы производства этой оборудование. За четыре десятилетия своего выпуска МТЗ-82.1 стал возделывать поля практически всех континентов нашей планеты. В частности, эти тракторы популярны в странах Азии, Африки и Южной Америки.
Основное назначение
Машина изначально создавалась для работы в поле. Хотя в принципе он достаточно многофункционален и представляет собой полноприводный средний тягач. Он вполне способен сеять различные зерновые культуры, собирать их, вспахивать поля, охлаждать. Очень часто трактор, оснащенный навесным оборудованием и прицепом, используется для перемещения различных грузов, а также дорожных или других земляных работ.
Некоторые нюансы
МТЗ-82.1 может эксплуатироваться практически в любых погодно-климатических условиях. По своей сути эта машина является точной копией (по функционалу) МТЗ-82. Единственным существенным отличием является лишь увеличенный объем салона модели с индексом 1.9.0003
В целом модель 82.1 как по внешним данным, так и по техническим характеристикам имеет вид полурамной конструкции, у которой задние ходовые колеса имеют значительно больший диаметр по сравнению с передними. Мотор, в свою очередь, расположен прямо под кабиной водителя.
Двигатель и коробка передач
Расход топлива МТЗ-82.1 совсем незначителен из-за наличия дизельного агрегата Д-243, который, как и трактор, также производится на Минском заводе. Мощность двигателя составляет 80 лошадиных сил, что позволяет машине ехать со скоростью 35 км/ч и одновременно использовать навесное оборудование для выполнения требуемой работы. Требуется минимальное техническое обслуживание.
Несмотря на то, что двигатель четырехтактный, на тракторе установлен электростартер, оборудованный предпусковым подогревателем. Также имеется жидкостная система охлаждения, которая позволяет запускать двигатель как в сильный мороз, так и в жару.
Что касается коробки передач, то до 1985 года этот трактор комплектовался исключительно трансмиссией механического типа, в которой число передач было не более 18×4 для передних колес и 16×4 для задних. В настоящее время задний мост можно заблокировать с помощью гидравлической системы. Надежная остановка дифференциала и его фиксация позволяют повысить проходимость техники.
Гидросистема
Включает в себя такие основные элементы:
- Гидронасос шестеренный типа НШ-32.
- Груз с гидравлическим захватом (используется для навесного оборудования).
- Силовые и позиционные регуляторы.
- Гидравлический цилиндр для прямого управления распределителем и рычажным механизмом.
Все эти узлы имеют возможность управления из кабины машиниста с помощью педалей и рычагов.
Основная суть гидрооборудования как новой модели, так и б/у трактора МТЗ-82. 1 заключается в высокоточной регулировке положения в пространстве используемого навесного оборудования на самых разнообразных типах грунтов и с различной подстилающей поверхностью.
Непосредственно на регуляторах расположены чувствительные датчики, реагирующие на любое изменение пространственного положения присоединяемого модуля и тяги. В целом гидросистема трактора позволяет увеличить производительность трактора и улучшить качество его работы (например, гарантировать равномерную вспашку земли с одинаковой глубиной).
Аппаратура
Новый МТЗ-82.1 комплектуется щитами с приборами, которые, в свою очередь, могут быть как автономными, так и входить в состав комбинированного оборудования. Кроме того, имеются блоки специальных сигнальных ламп и предохранитель с плавкими вставками для надежной защиты всех имеющихся электрических цепей.
Например, указатель температуры охлаждающей жидкости двигателя имеет сигнальные лампы разных цветов:
- Рабочий диапазон (от 80 до 100 градусов Цель) — зеленый.
- Нерабочий диапазон (до 80 градусов) — желтый.
- При температуре более 100 градусов Цельсия красный цвет.
Давление масла в дизеле должно быть в пределах 1-5 кгс/см 2 . Допускается скачок давления до 6 бар в момент пуска холодного двигателя. Однако, если в процессе дальнейшей работы аварийная лампа продолжает гореть, следует немедленно остановить двигатель и приступить к устранению неисправности. В свою очередь пневмосистема работает в диапазоне 5 — 8 кгс/см 2 и включает в себя компрессор и регулирующий клапан для управления дифференциальной работой тормозной системы.
Технические показатели
Любой трактор МТЗ-82.1 (б/у в т.ч.) имеет следующие технические данные:
- Мощность двигателя — 60 кВт.
- Номинальная скорость вращения — 2200 об/мин.
- Количество цилиндров 4 шт.
- Объем дизельного двигателя 4,75 л.
- Максимальный крутящий момент 290 Нм.
- Расход топлива при номинальной мощности — 220 г/кВт. ч.
- Запас по крутящему моменту составляет 15 %.
- Объем топливных баков 130 литров.
- Тип сцепления — однодисковое, сухое.
- Скорость движения вперед 1,89 — 33,4 км/ч.
- Скорость движения назад 3,98 — 8,97 км/ч.
- База станка 2450 мм.
- Длина — 3930 мм.
- Дорожный просвет под передним мостом — 645 мм.
- Просвет между дорогой и задней осью 465 мм.
- Эксплуатационная масса — 3900 кг.
Отзывы пользователей
Независимо от того, где был куплен б/у трактор МТЗ-82.1 (Авито, другие сайты или непосредственно у производителя), многие аграрии заметили, что производительность машины не слишком высока при обработке больших полей — более 80 га. Также мы отметили слабость третьей и шестой передач при больших нагрузках.
В свою очередь, двигатель не слишком чувствителен к качеству потребляемого топлива, однако при слишком низком качестве дизельного топлива двигатель может полностью заглохнуть или не запуститься. В этом случае, как показала практика, проблема устраняется регулировкой форсунок или заменой солярки.
Из положительных качеств трактора стоит отметить его практически абсолютную «неубиваемость», так как на него практически не действует ни жара, ни холод, ни бездорожье, ни пыль, ни любые возможные осадки. Кроме того, машина довольно легко сочетается со многими навесными приспособлениями и очень проста в эксплуатации. Водители также отмечают достаточно высокий уровень комфорта кабины, полностью соответствующий всем современным эргономическим нормам и требованиям.
Обкатка
Сегодня на Авито трактор 82.1 МТЗ можно приобрести абсолютно новым. Важно знать, что новая машина должна проходить неравномерную обкатку не менее 30 часов. Эта процедура является обязательной, поскольку позволяет всем без исключения частям трактора прижиться и обеспечить последующую долгую службу. Запрещается спускать с буксира новый, не раскатанный дизель во избежание быстрого износа его деталей.
Техническое обслуживание
Плановые работы проводятся по следующей схеме:
- Проверка натяжения ремня вентилятора (через 125 часов работы) — прогиб ремня не должен превышать 15 — 22 мм при нажатии с усилием 40 Н
- Регулятор люфта педали сцепления (после 500 часов работы) — 40 — 50 мм на ее подушке.
- Измерение люфта рулевого колеса (после 500 часов работы) — ни в коем случае не должен превышать 25% при работающем двигателе.
- Затяжка болтов ГБЦ (после 1000 часов работы) — момент затяжки должен быть в пределах 19 — 21 кгсм.
- Проверка затяжки гайки предохранительной муфты промежуточной опоры — муфта должна быть способна передавать крутящий момент 40 — 80 кгсм.
- Контроль уровня охлаждающей жидкости радиатора (через 10 часов работы трактора) — он должен быть не более чем на 50 мм ниже кромки заливной горловины.
- Замена масла в картере через 500 часов непрерывной работы.
- Замена фильтрующего элемента фильтра тонкой очистки — через 1000 часов непрерывной работы.
- Конденсат из ресивера сливается каждые десять часов работы.
- Оперативно переходите на подходящие сорта масла и смазки летом и зимой.
Заключение
Приобретая МТЗ-82.1 на Авито или в других местах, всегда помните, что этот трактор может активно использоваться как погрузчик, экскаватор, бульдозер и даже в некоторых случаях комбайн. А учитывая его относительно невысокую стоимость, становится понятно, почему спрос на него остается стабильно высоким на протяжении многих лет.
Mtz 82 Sebváltó — Alkatrészek
Használtakat is
Csak olcsók
Csak drágák
Házhozszállítás
Szűrés (Milyen sebváltó?):
- Összes sebváltó
Villa 095 sebváltó menesztő országuti terep ORIGINAL -50-1702095-A1-BY
Sebváltó
Arösszehasonlítás
4 320 FT
AgroRoll.hu
MTZ 82
Traktor
TRAZGERSIPUS 82
. 90-es évjárat Rendszáma és forgalmi engedélye nincs Üzemképes állapotú Ár: 1 100 000 Ft
1 100 000 Ft
mascus.hu
52-2308025 MTZ 82 NYOMTÁVÁLLÍTÓ HÜVELY
hüvely
Árösszehasonlítás
26 100 Ft
lokodiauto.hu
Belarus-MTZ 82 hektár harcos Póló ( L ) fekete — POLO-B-82-L
póló
Árösszehasonlítás
1 574 Ft
agroroll.hu
Mtz 50 -es váltó jó állapotban eladó
Használt
váltó
120 000 Ft
hasznaltat.hu
Eladó Mtz 80 panoráma
Használt
Eladó egy normál állapotú Mtz 80 1990-es évjáratú. Műszaki 2017.07-ig érvényes. Eladó MTZ 80 Panoráma
1 700 000 футов
MgApro. hu
Használt Mtz Váltó
Használt
Váltó
903 9023 553 553 5553 5553 555 5553 5555 555 555 555 555 555 555 555 5553 555 555 5553 555 5553 555 5553 553 553 553 553 553 553 553 553 553 553 553 553 553 553 5553 553 553 553 553 553 5.0223 MAN TGA-хозяин ZF16S2220TO Типовой номер Sebességváltó eladó.
Használt
Árösszehasonlítás
700 000 Ft
autoalkatresz.hu
MTZ 80 tractor (1984)
997 824 Ft
mascus.hu
MTZ 80
Traktor típus: Mezőgazdasági Traktor, Egyéb информация.: 80 LE 5 évig érvényes műszaki vizsga Üzemképes, jó állapotú Ar: 1 100 000 Ft ÁFA Érdeklődni …
1 100 000 Ft
MASCUS.HU
Renault Master Master 2. 5 Szívódízel Váltó, Indító, Gener …
152 400 FT
Autoalkatres.hu
vltta 5. www.motorvalto.hu alkatreszek auto-automotor-autovalto-alkatresz-1 … -29137 valto-525-tds-bmw 185181 Váltó 525 TDS BMW
30 000 Ft
Racing-bazar.hu
8 Ú 952.5 трактор (94 LE, CR, AdBlue, гар.)Traktor
Gyárilag beépített klímaberendezés új belarus traktorok magyarország egyik … új mtz 952.5 traktor (94 Le, Cr, Adblue, Gar.). 1025.2 kerekes traktor
traktor
5 924 581 Ft
autoline.hu
Fogaskerék MTZ-320 BELARUS ORIGINAL elsőhajtás — 320-2308026
fogaskerék
Árösszehasonlítás
12 000 Ft
agroroll.hu
Фогаскерек 198 II. meghajtó MTZ BELARUS ORIGINAL — 50-1701198-BY
meghajtó
Árösszehasonlítás
12 880 Ft
agroroll. hu
Fogaskerék MTZ-320 Z34 15 BELARUS ORIGINAL elsőhajtás — 320-2308018
fogaskerék
Árösszehasonlítás
8 500 Ft
agroroll.hu
Gebrauchte Беларусь МТЗ 1025 Турбо
12 788 Футов
landwirt.com
Беларусь МТЗ 1025 Турбо
12 788 футов
Landwirt.com
Belarus-Mtz 50 Póló (L) Fekete-Polo-B-50-L
Póló
árösszehasonlítás
1 574 FT
.hrorOllás
1 574 FT
.HROLOLL.HRROLLA
1 574 FT
.HROLOLL.HRROLLA
1 5744. ORIGINAL — 220-460805
szűrő
Árösszehasonlítás
13 800 Ft
agroroll. hu
Belarus MTZ 820
Új Belarus traktorok Magyarország egyik legnagyobb… Új Belarus MTZ 820 (80LE, Szöghajtás,garancia) Belarus MTZ 820
15 064 Ft
landwirt.com
Belarus MTZ 820.4 (egyenes hidas)
Új Belarus traktorok Magyarország egyik legnagyobb márkakereskedésétől…. Belarus MTZ 820.4 (egyenes hidas)
17 115 Ft
landwirt. Com
Belarus MTZ 920,3 Schlepper (2006)
Motor
MTZ 920,3 Schlepper, 2006, 90 PS, 4290 BTSD, Klimaanlage, Druckluftanlage, Turbo Motor, Guelarus Mtz 920.3 Schlepper.0003
landwirt.com
Беларусь МТЗ 920
Használt
2008-as évj., 700 üzemóra, jó állapotban eladó. 90Le Belarus MTZ 920
11 827 Ft
landwirt.com
Belarus MTZ 892. 2
készlet
5 650 000 Ft
landwirt.com
Belarus MTZ 550
Használt
Piros rendszám, felújított мотор, 90-ос гумик, жо аллапотбан эладо. Беларусь МТЗ 550
4 163 Ft
landwirt.com
Házi készítésű kis traktor
Használt
Házi készítésű kis traktor Opel Kadett Motor elsőkerék, hátsókerék hajtással, terepváltóval, kézi működtetésű hidraulikával 275000 Ft , Eke 20000 Ft …
275 000 Ft
mgapro. hu
Persely 191 fogaskerék 198 MTZ ORIGINAL — 50-1701191-BY
fogaskerék
Árösszehasonlítás
2 120 Ft
agroroll.hu
311 313 CDI 6 sebesseges valto, 2010 es автобол.
250 000 Ft
autoalkatresz. hu
MTZ 1221.3 tractor (2008)
Traktor típus: Mezőgazdasági Traktor, Kerékhajtás: 4 WD, Motorteljesítmény: 130, Vezetőfülke: Szokásos, Első kerékabroncs mérete: 420 70 R24, Hátsó …
6 142 854 Ft
mascus.hu
Mtz 50 генератор!
5 000 Ft
agrosat.hu
МТЗ кер кс лёк
5 000 Ft
agrosat.hu
MTZ 82.1
traktor
Traktor típus: Mezőgazdasági Traktor, Motorteljesítmény: 82
2 600 000 Ft
mascus.hu
MTZ 920.3 tractor (2006)
3 960 114 Ft
mascus.hu
MTZ 952.3 tractor (2007)
Traktor típus: Mezőgazdasági Traktor, Kerékhajtás: 4 WD, Motorteljesítmény: 90, Vezetőfülke: Szokásos, Első kerékabroncs mérete: 360 70 R24, Hátsó . ..
3 804 204 FT
MASCUS.HU
MTZ 82.1
HASSNált
EKE
TULAJDONOSTROOLól Aladó a Képen Látr5. , 1db pótkocsihúzó …
2 700 000 Ft
mgapro.hu
További sebváltó oldalak
- Használt MTZ váltó eladó
- MTZ 920 3 traktor kerékhajtás és váltó tesztelése
- Mtz traktor váltó
- Eladó váltó
- Mtz 50 váltó alkatrész
- Fiat ritmo váltó
- Váltó Zala megye
- Mtz váltó fogaskerék
- MTZ váltó tömítés
- MTZ SEBESSÉGVÁLTÓ ALKATRÉSZEK
- Mtz 80 váltó rajz
- Váltó
- John Deere bravúr DirectDrive sebességvalto
- sebességváltó ford mondeo 2 0 tdci Jármű
- sebességváltó javítás
- MTZ sebességváltó harom féle van
- mű9 Jezale Motors 82ki mű9 Jezale Motors 82153 Jezale Motors0154
- Sebességváltó egyben
- MTZ sebességváltó komplett 80 as
- Kirovec sebváltó próba Musica en tu Movil
- Kirovec sebváltó próba
- MTZ 82 traktor vezetéséről egy kis elméleti oktatás
- Mtz traktor alkatresz
- A BELARUS MTZ 50 és MTZ 52 traktorok
- MTZ Alkatrészek
- Mtz alkatrész rajz
- Belarus Mtz 82 kisfülkés
- MTZ Alkatrész
Sebváltó
Olcsó Mtz 82 Sebváltó MTZ SEBESSÉGVÁLTÓ ALKATRÉSZEK.
Master Renault Váltó Bowden
305×12 Renault Master -Hátsó — Renault Master
17 519 футов
Master, 2,5DCI, PF VALTOBOWNALY Váltóbovden, Európai, Olcsón. Opel Vivaro
130 FT
Renault Scenic II 1.9 DCI Manuál Sebváltó Váltó
50 000 футов
Renault Trafic Dízel 1995 CM
4 150 000 FT
- .0154
- Renault master sebességváltó bowden
- Kézifék bowden RENAULT MASTER III Box FV 2010
- Renault master váltó
- 8 Renault Master 2 5dci PF VÁltÓbowden váltóbovden
- RENAULT MASTER 2 5 DCI váltóbowden
Kapcsolódó lapok
- Renault 19 váltó 113
- Рено Валто 82
- Форд гэлакси автомата váltó 128
- Renault clio váltó 73
- Renault Twingo значение 86
- Renault Талия Валто 115
- Renault Trafic váltó 114
- Renault Megane váltó 109
- Автоматы Renault váltó 44
- Renault laguna váltó 88
- Renault сценический váltó 102
- Renault Clio Automatic váltó 89
- Renault сценический автомат váltó 99
- Renault Megane автомата váltó 109
- Renault laguna автомата значение 67
- Renault clio váltó szimering 117
- Renault master также gömbfej 95
- 6 sebességes váltó 116
- Ford Fiesta váltó 86
- Автомат Форд váltó 94
- Ford mondeo váltó 106
- Форд эскорт váltó 163
- Ford focus váltó 64
- Lada 5 sebességes váltó 86
- Simson 6 sebességes váltó 109
- Simson 4 sebességes váltó 144
- Мультикар váltó 88
- Опель цена 122
- Опель м32 цена 129
- Ford focus автомат váltó 125
- Ford focus Валто Боуден 76
- Ford mondeo автомата váltó 118
- Opel vectra b váltó 144
- Opel easytronic значение 63
- Опель виваро вальто 122
- Опель вектра цена 145
- Opel váltó szimering 138
- Opel astra f váltó 166
- Opel astra g váltó 131
- Opel corsa c váltó 144
- Автоматы Opel váltó 97
- Опель Корса Валто 139
- Опель Астра цена 137
- Opel vectra c автоматом váltó 176
- Opel omega автомата váltó 184
- Opel vectra auto váltó 175
- Renault master gyújtáskapcsoló 71
- Renault master fékbetét 158
- Renault master ajtózár 162
- Мастер Renault önindító 87
- Мастер Рено Альваз 58
- Renault мастер фельтенгели 98
- Renault master olajszűrő 160
- Renault master kormánymű 70
- Рено Мастер Фельни 139
- Renault мастер лапруго 82
- Рено мастер лего 87
- Renault master szélvédő 104
- Renault master vízhűtő 132
- Renault мастер улешузат 135
- Renault master тетоксомагтарто 122
- Рено мастер улес 126
- Renault мастер алькатрес 118
- Главный двигатель Renault 64
- Renault master экз. 146
- Renault master kuplungtárcsa 88
- Renault master ékszíjtárcsa 135
- Renault master hálófülke 47
- Рено мастер моторбурколат 86
- Renault master hidraulikus kinyomócsapágy 96
- Renault Master Hátsó féknyereg 139
- G dkg váltó 31
- Т4 вальто 50
- Уаз váltó 85
- Зф Валто 32
- Валто 146
- Мтз 82 Валто 50
- Dsg váltó 34
- Bmw smg váltó 56
- Мтз 50 вальто 90
- F13 значение 73
- Дуу Валто 73
- Кол-во см значение 67
- Фунты кг Валто 34
- M32 Валто 67
- Vw dsg váltó 65
- Асд Валто 61
- Фольксваген Валто 55
- МТБ Валто 217
Карбоангидразы как потенциальные мишени против дисфункции нервно-сосудистых единиц при болезни Альцгеймера и инсульте
Введение
Нейроваскулярная дисфункция является важным, ранним и причинным событием в патогенезе как болезни Альцгеймера (БА), так и инсульта (Iadecola, 2010, 2017; Hachinski et al. , 2019; Sweeney et al., 2019a; Freitas- Андраде и др., 2020; Злокович и др., 2020). Действительно, нейроваскулярная единица (НВУ) в последнее время набирает обороты в качестве фармакологической мишени при цереброваскулярных патологиях и нейродегенерации (Iadecola, 2017; Sweeney et al., 2019a; Zlokovic et al., 2020). NVU представляет собой функциональную многоклеточную структуру, состоящую из кровеносных сосудов и окружающих их различных типов клеток в центральной нервной системе (ЦНС), и играет важную роль в регуляции гомеостаза ЦНС (Andreone et al., 2015; Iadecola, 2017; Sweeney et al., 2019б; Фрейтас-Андраде и др., 2020). Важные функции NVU включают, среди прочего, регуляцию мозгового кровотока (CBF) и иммунологический надзор (Andreone et al., 2015; Cortes-Canteli and Iadecola, 2020; Freitas-Andrade et al., 2020). Дисфункция NVU наблюдается при старении, болезни Альцгеймера и после неврологических патологий, таких как инсульт (Iadecola, 2017; Cortes-Canteli and Iadecola, 2020; Freitas-Andrade et al. , 2020; Sarvari et al., 2020) и черепно-мозговая травма (Xing et al., 2012; Lok et al., 2015), среди прочих.
Инсульт и слабоумие являются двумя наиболее распространенными неврологическими расстройствами. Они создают риски друг для друга и имеют общие факторы риска, в основном поддающиеся изменению. Инсульт удваивает вероятность развития деменции (Savva and Stephan, 2010; Kuźma et al., 2018). Таким образом, профилактика инсульта путем лечения артериальной гипертензии и других факторов риска также может снизить заболеваемость деменцией (Kuźma et al., 2018; Hachinski et al., 2019).
AD является наиболее распространенной формой деменции и исторически характеризуется внеклеточными бета-амилоидными бляшками (Aβ) и внутриклеточными гиперфосфорилированными клубками тау-белка в определенных областях мозга (Day et al., 2015; Castillo-Carranza et al., 2017; Cortes). -Canteli and Iadecola, 2020; Ojo et al., 2021). Интересно, что было показано, что формы промежуточной агрегации Aβ и тау, такие как олигомеры, оказывают токсическое воздействие на несколько типов клеток NVU (Fossati et al. , 2010, 2012b; Parodi-Rullán et al., 2019).; Канепа и Фоссати, 2020 г.). Эта токсичность в сочетании с нарушением выведения нежелательного материала из мозга может привести к нейродегенерации и снижению когнитивных функций (Fossati et al., 2012b; Boland et al., 2018; Da Mesquita et al., 2018; Nortley et al. al., 2019; Provensi et al., 2019; Braun and Iliff, 2020; Carare et al., 2020; Cortes-Canteli and Iadecola, 2020; Nedergaard and Goldman, 2020; Parodi-Rullán et al., 2020; Quintana et al. др., 2021). Важно отметить, что до 90% пациентов с БА также имеют церебральную амилоидную ангиопатию (ЦАА), определяемую как отложение Aβ вокруг сосудов головного мозга и/или внутри стенок сосудов (Iadecola, 2017; Sweeney et al., 2019).а; Злокович и др., 2020). САА также часто встречается у пожилых людей без деменции и вносит важный вклад в дисфункцию NVU как при нормальном старении, так и при БА (Provensi et al., 2019; Cortes-Canteli and Iadecola, 2020; Ojo et al., 2021).
Инсульт классически определяется как неврологическое повреждение, связанное с острым очаговым повреждением ЦНС сосудистой причиной, включая инфаркт головного мозга, внутримозговое кровоизлияние (ВМК) и субарахноидальное кровоизлияние (САК), и является основной причиной инвалидности и смерти во всем мире (Sacco et al. , 2013). Наиболее распространенным типом инсульта является ишемический инсульт (ИИ), который возникает, когда атеросклеротические бляшки и жировые отложения вызывают окклюзию сосудов, прерывая кровоток в головном мозге. Кровеносным сосудом, который чаще всего закупоривается, является средняя мозговая артерия (Kuriakose and Xiao, 2020). Когда возникает ИИ, он сразу же вызывает множественные пагубные повреждения головного мозга из-за недостатка кислорода и глюкозы, а также связанной с этим провоспалительной сигнализации (Faraco et al., 2007; Freitas-Andrade et al., 2020; Kuriakose and Xiao). , 2020; Сарвари и др., 2020). После этого события наступает фаза реперфузионного повреждения, которая возникает при восстановлении кислорода и CBF (Faraco et al., 2007; Freitas-Andrade et al., 2020; Sarvari et al., 2020). Еще одним типом инсульта является геморрагический инсульт (ГС), который возникает при разрыве кровеносного сосуда, снабжающего кровью головной мозг (Corraini et al., 2017; Sarvari et al. , 2020). HS характеризуется большим объемом поражения, более высоким внутричерепным давлением и вызывает более тяжелое повреждение головного мозга, чем ИИ. Важно отметить, что ИИ и ГС влияют на разные области мозга (Corraini et al., 2017).
Общими факторами риска БА и инсульта являются снижение мозгового кровообращения, сердечно-сосудистые заболевания и возраст (Sarvari et al., 2020; Ojo et al., 2021). Факторы сердечно-сосудистого риска, такие как ожирение, диабет, гипертония и атеросклероз, усугубляют цереброваскулярную патологию, а также нейродегенерацию, включая болезнь Альцгеймера (Girouard and Iadecola, 2006; de Bruijn and Ikram, 2014; Cortes-Canteli and Iadecola, 2020; Sarvari et al. ., 2020). Общие механизмы, лежащие в основе как БА, так и инсульта, включают нейровоспаление, митохондриальную дисфункцию, гибель клеток и нарушение регуляции гематоэнцефалического барьера (ГЭБ), что указывает на то, что NVU является мишенью для обоих заболеваний (Fossati et al., 2012b; Alluri et al. , 2014; Секердаг и др., 2018; Эльдахшан и др., 2019.; Провенси и др., 2019; Пароди-Руллан и др., 2020). Огромное количество исследований было потрачено на открытие новых методов лечения атопического дерматита и инсульта. При БА это недавно привело к неоднозначному одобрению FDA адуканумаба (Ferrero et al., 2016; Sevigny et al., 2017; Knopman et al., 2021). Тем не менее, необходимо провести дополнительные исследования, чтобы разработать успешную модифицирующую болезнь терапию для обоих расстройств. Научное сообщество особенно поощряет изучение перепрофилированных препаратов, одобренных FDA для лечения других заболеваний, которые могут быть полезны при болезни Альцгеймера и инсульте, позволяя при этом более быстро перевести их в клинические испытания.
В этом обзоре будет представлена идея потенциального перепрофилирования ингибиторов карбоангидразы (CAI), многие из которых уже одобрены FDA для других показаний, для профилактики цереброваскулярной и нейроваскулярной патологии при AD и инсульте, а также подчеркнуто влияние карбоангидразы (CAI). ) модуляция этих двух доминирующих неврологических расстройств.
КА представляют собой семейство металлоферментов цинка, которые катализируют обратимую гидратацию диоксида углерода с образованием бикарбоната и протона (CO 2 + H 2 O ↔ HCO 3 – + H + ) (Supuran, 2011; Mishra et al., 2020). Эта химическая реакция необходима для многих физиологических процессов, таких как pH и гомеостаз ионов, транспорт углекислого газа, секреция электролитов, глюконеогенез, липогенез и уреагенез, реабсорбция воды и натрия в почках, реабсорбция и кальцификация костей, образование и оборот спинномозговой жидкости, среди прочих процессов (Provensi et al., 2019; Zamanova et al., 2019; Мишра и др., 2020). КА были изучены как хорошо известная фармакологическая мишень для многих периферических расстройств и расстройств ЦНС (Bradwell et al., 1992; Ilies et al., 2004; De Simone and Supuran, 2007; Supuran, 2008, 2018; Akocak and Ilies, 2014). ; Провенси и др. , 2019; Заманова и др., 2019; Мишра и др., 2020). Интересно, что многие недавние исследования выявили общую цель дальнейшего выяснения участия КА как в БА, так и в инсульте.
У человека есть 15 изоформ КА, все с разными паттернами экспрессии на тканевом и клеточном уровне (Supuran, 2011; Provensi et al., 2019).; Заманова и др., 2019; Мишра и др., 2020). Многие из этих изоформ экспрессируются в NVU, и было высказано предположение, что каждая из них участвует в различных функциях (Draghici et al., 2014; Rasmussen and Boedtkjer, 2018; Provensi et al., 2019; Zamanova et al., 2019). Известно, что одни изоформы являются внеклеточными, закрепленными на плазматической мембране (СА-IV, СА-IX и СА-XII, СА-XIV), а другие — цитозольными (СА-I, СА-II, СА-III, CA-VII, CA-XIII), две находятся в митохондриях (CA-VA и CA-VB) (Boriack-Sjodin et al., 19).95; Nishimori et al., 2005), некоторые из них представляют собой акаталитические изоформы (CA-VIII, CA-X и CA-XI) (Aspatwar et al., 2014), а одна изоформа секретируется в слюну (CA-VI) (Nishimori et al. al., 2007; Alterio et al., 2012; Patrikainen et al., 2014; Provensi et al., 2019; Zamanova et al., 2019; Mishra et al., 2020). Функция СА была связана с патологией БА, а также с инсультом, но конкретные изоформы и вовлеченные патологические механизмы до конца не изучены (Wang et al., 2009; Draghici et al., 2014; Fossati et al., 2016; Pollard et al. и др., 2016; Солесио и др., 2018; Мишра и др., 2020). Определение роли каждой изоформы КА в норме и заболевании должно быть приоритетом для разработки новых и эффективных методов лечения цереброваскулярной патологии (Provensi et al., 2019).; Мишра и др., 2020).
ИАИ сначала были разработаны как мочегонные средства и стали эффективными при лечении глаукомы, отека мозга, эпилепсии, а также высотной болезни (Mincione et al., 2007; Supuran, 2008; Ritchie et al., 2012; Akocak and Ilies, 2014; Заманова и др., 2019; Мишра и др., 2020). Одобренные FDA CAI метазоламид (MTZ) и ацетазоламид (ATZ) являются наиболее изученными пан-CAI. Их деятельность в NVU будет одним из основных направлений этого обзора. Другие CAI, одобренные FDA, такие как топирамат, обладающий некоторой селективностью в отношении митохондриальных изоформ CA-VA и CA-VB, наряду с соединениями, обладающими селективностью в отношении CA-IX и CA-XII, также будут обсуждаться (Scozzafava et al., 2000; Supuran, 2012; Andring et al., 2020; McDonald et al., 2020). Было обнаружено, что МТЗ и АТЗ, наряду с топираматом, обладают защитными свойствами в отношении цереброваскулярной патологии, а также в отношении функции митохондрий, важной мишени для целостности НВУ (Wang et al., 2009).; Прайс и др., 2012; Фоссати и др., 2016; Солесио и др., 2018 г.; Саламе и др., 2019). Значительное количество литературы по одобренным FDA пан-CAI указывает на то, что они безопасны и могут пройти BBB. Однако разработка новых соединений, нацеленных на конкретные изоформы, может повысить эффективность и уменьшить побочные эффекты уже существующих пан-ЦАИ (Provensi et al., 2019; Mishra et al., 2020).
Этот обзор сначала проиллюстрирует базовую структуру и функции клеток, составляющих NVU, в частности, описав, как они становятся дисфункциональными при инсульте и болезни Альцгеймера. Затем мы обсудим свойства нескольких изоформ КА и выделим имеющиеся данные, показывающие, как ингибирование КА может быть защитным от множественных нарушений регуляции в клетках, составляющих NVU, указывая на КА как на потенциальную мишень для терапии как инсульта, так и БА (Provensi et al., 2019; Мишра и др., 2020).
Нейроваскулярная единица: функция и дисфункция при инсульте и болезни Альцгеймера
Нейроваскулярная единица
Типы клеток, которые составляют NVU (изображены на рис. 1) и взаимодействуют для выполнения своих функций, включают эндотелиальные клетки (ЭК), перициты, гладкие мышечные клетки (ГМК), астроциты и микроглия (Iadecola, 2017; Freitas-Andrade et al., 2020), которые функционально или физически связаны с нейронами (Andreone et al., 2015; Cortes-Canteli and Iadecola, 2020; Freitas -Андраде и др., 2020). NVU представляет собой морфофункциональную единицу, включающую ГЭБ, которая важна для транспорта питательных веществ и кислорода из большого круга кровообращения в мозг, для выведения токсических отходов из ЦНС, для связи между кровотоком и функцией нейронов, а также для формирования физического барьера для предотвращения проникновения патогенов и других вредоносных объектов в ЦНС (Sarvari et al. , 2020).
Рис. 1. Сосудисто-нервный блок. Рисунок, изображающий типы клеток, которые составляют NVU в капиллярах головного мозга. Капилляры головного мозга окружены перицитами, как показано на этом рисунке, а артерии и артериолы окружены ГМК. Другими важными клетками, связанными с кровеносными сосудами и важными для ГЭБ и нейроваскулярных функций, также представленными на этом рисунке, являются астроциты и микроглия. ECs, эндотелиальные клетки; ГМК, гладкомышечные клетки; ГЭБ, гематоэнцефалический барьер; NVU, сосудисто-нервная единица; ТК, клетки микроглии, КМ, базальная мембрана.
Эндотелиальные клетки
ЭК являются важными компонентами стенки кровеносного сосуда. Цереброваскулярные ЭК образуют плотные и спаянные соединения друг с другом, чтобы ограничить поступление молекул и клеток из периферического кровообращения в ЦНС (Reese and Karnovsky, 1967). Транспортеры, экспрессируемые на плазматической мембране ЭК, специфически регулируют то, что входит и выходит из ЦНС. Например, глюкоза поступает в мозг исключительно через транспортеры, несмотря на то, что мозг является высокометаболическим органом, ответственным за до 25% общего потребления глюкозы в организме (Tang et al., 2017; Parodi-Rullán et al., 2019).). ЭК также регулируют кровоток, высвобождая вазодилататоры и вазоконстрикторы, такие как оксид азота (NO) и эндотелин-1 соответственно (Morikawa et al., 1994; Biernaskie et al., 2001; Freitas-Andrade et al., 2020). В связи с тем, что митохондрии очень многочисленны в цереброваскулярных ЭК (Oldendorf et al., 1977; Sarvari et al., 2020), эти клетки особенно чувствительны к кислородному голоданию (Pun et al., 2009; Freitas-Andrade et al. , 2020). Таким образом, патологические состояния, вызывающие кислородно-глюкозную депривацию (ОГД) и вызывающие избыточную выработку активных форм кислорода (АФК), вызывают дисфункцию церебрального эндотелия, гибель клеток и разрушение ГЭБ (Schreibelt et al., 2007; Pun et al., 2009).; Лоххед и др., 2010; Гизо и др. , 2014; Фоссати и др., 2016; Пароди-Руллан и др., 2019 г.; Freitas-Andrade et al., 2020), указывая на митохондрии как на критически важные мишени для функции EC и целостности ГЭБ. Эффективная коммуникация и обмен материалами между ECs и другими типами клеток NVU необходимы для гомеостаза ЦНС (Andreone et al., 2015; Freitas-Andrade et al., 2020; Sarvari et al., 2020).
Цереброваскулярная дисфункция наблюдается как при БА, так и при ИИ (Fossati et al., 2010, 2012a,b; Guo et al., 2010; Lochhead et al., 2010; Jiao et al., 2011; Shin et al., 2016 г.; Фрейтас-Андраде и др., 2020 г.; Пароди-Руллан и др., 2020 г.; Кинтана и др., 2021 г.). Старение, а также сердечно-сосудистые факторы риска, такие как гипертония и диабет, способствуют развитию цереброваскулярной патологии (Huang et al., 19).95; Жируар и Ядекола, 2006 г.; Прайс и др., 2012; Кортес-Кантели и Ядекола, 2020 г.; Оджо и др., 2021). Снижение количества белков плотного контакта, таких как зона окклюдина-1 (ZO-1) и окклюдин, наблюдается на моделях инсульта и атопического дерматита (Marco and Skaper, 2006; Jiao et al. , 2011; Engelhardt et al., 2014; Freitas- Andrade et al., 2020; Parodi-Rullán et al., 2020). Повышение экспрессии молекул адгезии, таких как молекула межклеточной адгезии-1 (ICAM-1) и молекула адгезии сосудистых клеток-1 (VCAM-1), наблюдается при инсульте, а в последнее время также было связано с атопическим дерматитом (Cruz Hernandez et al. др., 2019; Сарвари и др., 2020). Уменьшение белков плотного контакта и увеличение количества молекул адгезии запускает рекрутирование периферических иммунных клеток в головной мозг и может быть связано с активацией эндотелия молекулярными паттернами, связанными с опасностью (DAMP), такими как низкий уровень глюкозы и кислорода, и агрегированными белками, такими как Aβ и гиперфосфорилированный тау (Canepa and Fossati, 2020; Freitas-Andrade et al., 2020; Sarvari et al., 2020). Увеличение проницаемости ГЭБ позволяет периферическим веществам проникать в ЦНС, что приводит к нейровоспалению и окислительному стрессу (Pun et al., 2009).; Тернер и Шарп, 2016 г. ; Ян и др., 2019). Повышенный окислительный стресс в ЭК усугубляет митохондриальную дисфункцию, приводя к апоптозу (Lochhead et al., 2010; Fossati et al., 2012b, 2016; Solesio et al., 2018; Parodi-Rullán et al., 2019). Как во время ишемического повреждения, так и при АтД ЭК снижают выработку NO, вызывая нарушение регуляции CBF (Morikawa et al., 1994; Biernaskie et al., 2001; Girouard and Iadecola, 2006; Austin et al., 2013; Parodi- Rullán et al., 2019; Freitas-Andrade et al., 2020). Интересно, что у мышей с нокаутом (KO) эндотелиальной синтазы оксида азота (e-NOS) дефицит NO снижает способность ECs нейтрализовать ROS, усиливая окислительный стресс и нейровоспаление, что, вероятно, усугубляет патологию AD (Austin et al., 2013). Соответственно, в моделях БА и инсульта было показано, что дисфункция ЭК усиливает нейровоспаление за счет увеличения продукции АФК и повышения проницаемости ГЭБ среди других клеточных механизмов (Parodi-Rullán et al., 2019)., 2020; Ян и др., 2019).
Перициты и гладкомышечные клетки
Перициты и ГМК окружают ЭК внутри сосудистых стенок, охватывая капилляры и артериолы/артерии, соответственно. Оба типа клеток играют жизненно важную роль в регуляции целостности CBF и BBB (Sagare et al., 2013; Hall et al., 2014; Freitas-Andrade et al., 2020). Эти клетки регулируют кровоток в основном за счет своего сокращения или расширения, которые контролируют диаметр кровеносного сосуда (Hall et al., 2014; Freitas-Andrade et al., 2020). В последнее время тщательно исследована роль кровеносных сосудов в клиренсе интерстициальной жидкости и белков ЦНС (Aldea et al., 2019).; Караре и др., 2020). Было высказано предположение, что интрамуральный периартериальный дренаж (IPAD) представляет собой путь оттока мозговой интерстициальной жидкости и растворимых белков из ЦНС в шейные лимфатические узлы путем прохождения по капиллярам и артериолам в направлении, противоположном кровотоку, в пределах базальной мембраны шейных позвонков. сосудистая сеть головного мозга. Сокращение SMC обеспечивает движущую силу для IPAD (Carare et al., 2008; Aldea et al., 2019). Эта система не регулируется в моделях ЦАА и старения (Ojo et al. , 2021). Другой путь очистки головного мозга, глимфатическая система, наблюдался для импорта спинномозговой жидкости по периартериальному пространству и экспорта интерстициальной жидкости по перивенозным пространствам в периваскулярных туннелях, образованных астроглиальными клетками. Было высказано предположение, что этот путь играет важную роль в клиренсе и регуляции токсичных белков в головном мозге, таких как тау и Aβ, и опосредован астроцитарным аквапорином-4 (AQP4) (Braun and Iliff, 2020; Nedergaard and Goldman, 2020) . Интересно, что глимфатическая система наиболее эффективна во время сна (Boespflug and Iliff, 2018; Mestre et al., 2020).
Как во время инсульта, так и при БА наблюдается отсоединение перицитов от ГЭБ (Sagare et al., 2013; Freitas-Andrade et al., 2020). В мышиной модели AD потеря перицитов усугубляла патологию AD (Sagare et al., 2013). Потеря перицитов снижает сосудистую стабильность и снижает способность NVU регулировать CBF (Sagare et al., 2013; Hall et al. , 2014). С другой стороны, недавнее исследование пришло к выводу, что Aβ индуцирует опосредованное перицитами сужение капилляров, снижая CBF (Nortley et al., 2019).). Было замечено, что перициты повреждаются в моделях диабета, а также ожирения, которые являются факторами риска AD и IS (Price et al., 2012; Shah et al., 2013a). У кошек, подвергшихся гипоксии, перициты отделялись от микроциркуляторного русла (Gonul et al., 2002). Наряду с нарушением регуляции CBF в моделях БА и инсульта потеря перицитов и ГМК также способствует нарушению клиренса в моделях БА и ЦАА (Sagare et al., 2013; Aldea et al., 2019; Carare et al., 2020; Kim и др., 2020; Оджо и др., 2021).
Астроциты
Астроциты являются важными клетками для NVU и обеспечивают физическую связь между кровеносными сосудами и нейронами (Iadecola and Nedergaard, 2007; Braun and Iliff, 2020). Концы астроцитов оборачивают кровеносные сосуды, помогая стабилизировать плотные соединения ЭК и регулировать CBF (Iadecola and Nedergaard, 2007). Астроциты играют ключевую роль в обеспечении метаболической и физической поддержки ЦНС, наряду с регуляцией CBF, а также очисткой мозга (Freitas-Andrade et al., 2020). AQP4 представляет собой мембранный белок, участвующий в водном обмене в ЦНС. У здоровых людей AQP4 представляет собой мембранный канал, локализованный на концах астроцитов, и он важен для выведения токсичных растворенных веществ из мозга (Smith et al., 2019).). Предполагается, что AQP4 является частью глимфатической системы, опосредуя обмен жидкости и дренаж белков, а также выведение жидкости из ЦНС (Iliff et al., 2012; Boland et al., 2018; Braun and Iliff). , 2020; Местре и др., 2020). Несмотря на то, что дифференциальный вклад глимфатической системы и пути IPAD в клиренс мозговой жидкости и отходов не полностью изучен, считается, что астроциты играют ключевую роль в обмене жидкостей, основываясь на исследованиях, показывающих, что в AQP4 KO мышей, приток спинномозговой жидкости, а также клиренс ЦНС были снижены (Iliff et al. , 2012; Braun and Iliff, 2020). Другой важной функцией астроцитов является поглощение и высвобождение глутамата, что очень важно для поддержания гомеостаза ЦНС (Dejakaisaya et al., 2021; van Putten et al., 2021; Verkerke et al., 2021). Астроциты также играют решающую роль в поддержании антиоксидантной системы мозга и в производстве глутатиона, важного модулятора окислительного стресса и старения (Bains and Shaw, 19).97; Венкатешаппа и др., 2012 г.; Ховарт и др., 2017 г.; Веркерке и др., 2021).
При БА астроциты теряют свою поляризацию, отсоединяются от ГЭБ, становятся реактивными и выделяют воспалительные цитокины (Liddelow et al., 2017; Sweeney et al., 2019b; Cortes-Canteli and Iadecola, 2020). Перекрестные помехи между астроцитами и микроглией нуждаются в дальнейшем понимании, хотя было широко показано, что они модулируют состояние активации друг друга (Joshi et al., 2019; McConnell et al., 2019; McAlpine et al., 2021). Сообщалось также, что астроциты теряют экспрессию AQP4 на ранних стадиях болезни Альцгеймера (Smith et al. , 2019).). Иными словами, при инсульте экспрессия AQP4 коррелирует с отеком головного мозга, увеличивая повреждение нейронов (Manley et al., 2000). При ИИ астроглиоз очень важен из-за присутствия DAMP в начальной/острой фазе, а также во вторичной/поздней фазе повреждения. Основным показателем глиоза является увеличение экспрессии глиального фибриллярного кислого белка (GFAP) астроцитами, наблюдаемое как при инсульте, так и в моделях болезни Альцгеймера (Pekny and Lane, 2007; Oeckl et al., 2019; Freitas-Andrade et al. , 2020). При ишемическом повреждении нервно-сосудистая связь теряется вместе с опосредованным астроцитами CBF (McConnell et al., 2019).; Фрейтас-Андраде и др., 2020). Это приводит к циклу метаболического стресса и воспаления, усугублению митохондриальной дисфункции в сосудистых клетках и, в конечном итоге, к разрушению ГЭБ (Girouard and Iadecola, 2006; Freitas-Andrade et al., 2020; Radenovic et al., 2020; Sarvari et al. ., 2020).
Микроглия
Микроглия, хотя и не прикреплена напрямую к ЭК, обеспечивает целостность ГЭБ (Eldahshan et al. , 2019; Freitas-Andrade et al., 2020). Они считаются резидентными иммунными клетками ЦНС, способными фагоцитировать нейротоксические вещества, а совсем недавно сообщалось, что они также участвуют в ряде церебральных гомеостатических функций, включая синаптическую пластичность и развитие мозга (Badimon et al., 2020). ; Уиллис и др., 2020). Существует два хорошо изученных фенотипа активации микроглии, обозначаемые как M1 и M2, которые классифицируются как провоспалительные и противовоспалительные соответственно. Фенотип M1 вреден для ГЭБ, вызывая нарушение регуляции NVU, в то время как M2 способствует эндоцитозу и клиренсу токсических веществ и отмирающих клеток, ограничивая количество окислительного стресса и создавая более подходящую среду для регенерации/заживления после ЦНС. травмы (Танг и Ле, 2016; Фрейтас-Андраде и др., 2020; Цзян и др., 2020). Недавно было обнаружено, что белок, высоко экспрессируемый в микроглии, запускающий рецептор, экспрессируемый на миелоидных клетках-2 (TREM2), защищает от нейродегенерации в моделях AD. Предполагается, что TREM2 обладает нейропротекторным действием, связанным с фенотипом М2, хотя существуют варианты, повышающие вероятность развития болезни Альцгеймера (Ulland and Colonna, 2018; Gervois and Lambrichts, 2019).; Чжоу и др., 2020). Защитные механизмы TREM2 также наблюдались при ИИ (Gervois, Lambrichts, 2019).
После ИИ или СГ микроглия переходит в реактивное состояние, вызывая высвобождение провоспалительных цитокинов, усугубляя нейровоспаление и нейроваскулярную дисфункцию (Eldahshan et al., 2019). Активация микроглии также наблюдается в моделях AD и в настоящее время считается одним из основных признаков заболевания (Villegas-Llerena et al., 2016). Активация воспаления наблюдается при старении, нейродегенерации, а также при инсульте (Mohamed et al., 2015; Liddelow et al., 2017; Hu et al., 2019).; Ян и др., 2019). В головном мозге активация воспаления происходит во многих различных типах клеток. В частности, микроглия, будучи наиболее чувствительными клетками к патоген-ассоциированным молекулярным паттернам (PAMP) и DAMP, запускает активацию каспазы-1 и высвобождение цитокинов IL-1β вместе с IL-18 (Hu et al. , 2019). Затем активируются цитокиновые рецепторы на сосудистых клетках, запуская вредные процессы, которые вызывают дисфункцию NVU и нарушение нейроваскулярной связи (Eldahshan et al., 2019).). Среди других клеток микроглия также секретирует металлопротеазы (ММР), такие как матриксная металлопептидаза-2 (ММР-2) и матриксная металлопептидаза-9 (ММР-9), которые, как сообщается, активируются при БА и ИИ. разрушая базальную мембрану, тем самым увеличивая проницаемость ГЭБ и дисфункцию НВУ (Lorenzl et al., 2003; Hernandez-Guillamon et al., 2010; Yang and Rosenberg, 2015; Turner and Sharp, 2016; Montaner et al., 2019; Freitas- Andrade et al., 2020; Sarvari et al., 2020; Carcel-Marquez et al., 2021).
Базальная мембрана
Базальная мембрана является важной частью ГЭБ и состоит из многочисленных белков, таких как ламинины, коллаген, нидоген и протеогликаны сульфата гепарина (Carare et al., 2020). Различные белки, составляющие базальную мембрану, секретируются типами клеток, составляющими NVU, такими как ECs, перициты и астроциты. Эти разные белки поддерживают межклеточные взаимодействия и, таким образом, целостность ГЭБ (Carare et al., 2020; Freitas-Andrade et al., 2020).
Разрушение базальной мембраны резко проявляется после окклюзии средней мозговой артерии (MCAO) (Sarvari et al., 2020). Эта дегенерация происходит за счет различных механизмов нейровоспаления, таких как продукция АФК, а также секреция ММР (Mohamed et al., 2015; Yao, 2019; Freitas-Andrade et al., 2020). Кроме того, секреция и активация ММП нарушают состав ГЭБ, вызывая его негерметичность и еще больше усугубляя уже существующие окислительный стресс и нейровоспаление (Fujimura et al., 19).99; Эрнандес-Гильямон и др., 2010 г.; Монтанер и др., 2019 г.; Кан и Яо, 2020 г.). В моделях CAA также наблюдалось изменение состава и структуры базальной мембраны (Morris et al., 2014).
Нейроны
Нейроны, связанные с кровеносными сосудами, связываются с ECs и другими клетками NVU для модуляции сосудистой структуры, расширения или сужения, а также для обеспечения питательными веществами в зависимости от потребности нейронов (Andreone et al. , 2015; Iadecola, 2017; Kaplan и др., 2020). Нейроваскулярная связь необходима для поддержания надлежащего притока питательных веществ и надлежащего оттока токсичных отходов из мозга (Girouard and Iadecola, 2006; Andreone et al., 2015; Kaplan et al., 2020). Было обнаружено, что активность нейронов участвует как в ангиогенезе, так и в нейроваскулярной связи (Girouard and Iadecola, 2006; Huneau et al., 2015).
При повреждении ЦНС и гибели клеток NVU происходит нарушение регуляции нейроваскулярной связи (Iadecola, 2017; Sweeney et al., 2019a; Freitas-Andrade et al., 2020; Kaplan et al., 2020). Отсутствие связи между ЦНС и системным кровообращением в конечном итоге приводит к метаболическому сбою, окислительному стрессу, митохондриальной дисфункции и синаптической потере (Freitas-Andrade et al., 2020; Kaplan et al., 2020). Таким образом, возможно, что при сохранении целостности и функции сосудов головного мозга нейродегенерация протекает менее тяжело и лучше поддерживается гомеостаз ЦНС.
Карбоангидразы
КА представляют собой семейство металлоферментов цинка, катализирующих обратимую реакцию CO 2 + H 2 O ⇌ HCO 3 – + H + . Все четыре вида в этой химической реакции необходимы для гомеостаза ЦНС. Поэтому ожидается, что СА влияют на ЦНС и дальнейшую функцию НВУ. Реакция гидратации CO 2 может протекать спонтанно (не катализируемая), но скорость реакции слишком мала для динамики живых клеток. Следовательно, КА повсеместно распространен во всех живых организмах, в том числе и в человеке, где ускоряет эту реакцию в миллионы раз, делая взаимопревращение СО 2 и HCO 3 – почти мгновенно. У человека существует 15 изоформ СА, либо акаталитических (СА-VIII, СА-X и СА-XI), точная функция которых в настоящее время неизвестна, либо каталитически активных. Последние находятся в субклеточных локациях, где требуется взаимопревращение СО 2 /HCO 3 – , из места продукции в митохондриях (митохондриальные КА-ВА и КА-ВБ), продвигаясь в цитозоль (КА -I, СА-II, СА-III, СА-VII, СА-XIII), затем на плазматическую мембрану (СА-IV, СА-IX, СА-XII, СА-XIV) и, наконец, внеклеточно (СА-VI секретируется). в слюне (Супуран, 2008; Заманова и др., 2019; Мишра и др., 2020). В качестве генетических манипуляций и селективных ингибиторов (Supuran et al., 1998; Ilies et al., 2003; Smaine et al., 2007; Supuran, 2008; Güzel et al., 2009; Winum et al., 2009; Alterio et al. , 2012; Akocak et al., 2016; Angeli et al., 2020) стали более доступными, увеличилось количество исследований по определению клеточно-специфической функции различных изоформ КА (Ghandour et al., 1992; Sly and Hu, 1995; Kida et al., 2006; Pan et al., 2006, 2012; Imtaiyaz Hassan et al., 2013; Shah et al., 2013b; Akocak and Ilies, 2014; Angeli et al., 2017; Waheed и Sly, 2017; Хаапасало и др., 2020; Мишра и др., 2020). Одной из наиболее широко экспрессируемых и каталитически активных изоформ является CA-II, имеющая скорость оборота CO 2 гидратация приближается к пределу диффузии (K cat = 1,4 × 10 6 с –1 ). Это цитозольный фермент, который имеет самое широкое распространение в организме человека, экспрессируясь в клетках практически каждой ткани или органа. В головном мозге он содержится в больших количествах в олигодендроцитах и эпителии сосудистых сплетений. Субъекты, страдающие синдромом дефицита CA-II, аутосомно-рецессивным заболеванием человека, демонстрируют остеопетроз, хрупкость костей, ацидоз почечных канальцев и, что важно, церебральную кальцификацию и когнитивные дефекты, задержку развития и, как правило, низкий рост (Sly and Hu, 19).95; Супуран, 2008 г.; Заманова и др., 2019; Мишра и др., 2020). Было обнаружено, что эта изоформа перемещается в митохондрии при старении и нейродегенерации на мышиной модели дегенерации клеток Пуркинье (Pollard et al., 2016). Это исследование также показало, что C. elegans , подвергшиеся воздействию CA-II, имеют более короткую продолжительность жизни, что позволяет предположить, что высокие уровни CA-II участвуют в механизмах, ограничивающих жизненный цикл/цикл клеток. Будущие исследования должны дополнительно выяснить, почему и как это происходит.
Изоформа СА-I впервые была идентифицирована в эритроцитах, где ее в пять-шесть раз больше, чем СА-II, хотя на нее приходится лишь около 15% активности СА-II (Supuran, 2008; Zamanova et al. , 2019; Мишра и др., 2020). CA-I является наиболее распространенным негемоглобиновым белком в эритроцитах и вместе с CA-II способствует уравновешиванию пулов растворенного CO 2 /HCO 3 – в крови и поддерживает гомеостаз pH крови, а также способствует транспорт CO 2 из головного мозга и метаболизирующих тканей в легкие (Sly and Hu, 1995; Supuran, 2008; Zamanova et al., 2019; Mishra et al., 2020). Предполагается, что CA-I способствует отеку головного мозга из-за наблюдения, что CA-I увеличивается в мозге после HS (Guo et al., 2012). Одним из наиболее важных цитозольных изоферментов в головном мозге является CA-VII, быстрый изофермент (K cat = 9,5 × 10 5 с –1 ), обнаруживаемый преимущественно в нейронах гиппокампа вместе с CA-II. Интересно, что CA-VII не обнаружен в глиальных клетках, которые содержат только CA-II в цитозоле. Помимо ЦНС, CA-VII обнаруживается в скелетных мышцах, желудке, двенадцатиперстной кишке, печени, толстой кишке (Sly, Hu, 1995; Supuran, 2008; Zamanova et al. , 2019; Mishra et al., 2020). В мозге группа Кайлы показала, что CA-VII действует как молекулярный переключатель в пирамидных нейронах CA1 гиппокампа при развитии синхронного гамма-излучения в ответ на высокочастотную стимуляцию. Это открытие делает CA-VII важным модулятором долговременной потенциации, синаптической пластичности, памяти и процессов обучения (Ruusuvuori et al., 2004). Та же группа, использующая новый CA-VII ( Car7 ) KO мышиная модель, а также CA-II ( Car2 ) KO и CA-II/VII двойные KO мышиные модели показали, что в зрелых пирамидных нейронах гиппокампа CA-VII и CA-II изоферменты усиливают бикарбонат-управляемое ГАМКергическое возбуждение во время интенсивной активации ГАМК A -рецепторов. Экспрессия этих двух цитозольных изоферментов была обнаружена в очень раннем возрасте у животных (через 10 и 20 дней после рождения), что указывает на то, что СА-VII и СА-II являются ключевыми молекулами в возрастной регуляции рН нейронов (Supuran , 2008; Руусувуори и др. , 2013; Руусувуори и Кайла, 2014; Заманова и др., 2019.; Мишра и др., 2020).
Связанные с мембраной изоферменты также играют важную роль в мозге, особенно в регуляции внеклеточного pH (Chesler, 2003; Shah et al., 2005). Так, фосфатидилинозитолгликановый (GPI) заякоренный изофермент СА-IV (Stams et al., 1996) представляет собой быстрый изофермент (K cat = 1,1 × 10 6 с –1 ), обнаруживаемый в плазме крови. поверхность корковых капилляров. Он более устойчив к ингибированию ионами галогенидов, чем CA-II, поскольку он адаптирован для выполнения CO 2 /HCO 3 – взаимопревращение во внеклеточном пространстве, которое содержит более высокую концентрацию ионов Cl – , чем цитозоль. Изофермент также экспрессируется в хориокапиллярах глаза, в скелетных и сердечных мышцах, легких, почках, желудочно-кишечном и репродуктивном трактах (Sly, Hu, 1995; Supuran, 2008; Zamanova et al., 2019; Mishra et al. , 2020). ). Другой изоформой, локализованной в плазматической мембране, является СА-IX, трансмембранный изофермент, обладающий N-концевым протеогликановым доменом, каталитическим доменом, однопроходной трансмембранной областью и внутриклеточным хвостом. Это димерный белок с низким паттерном экспрессии в большинстве органов, за исключением пищеварительной системы (наибольшее количество) и ЦНС, где его можно обнаружить главным образом в клетках, выстилающих желудочки, и в сосудистом сплетении (Saarnio et al., 19).98; Хилво и др., 2008 г.; Супуран, 2008 г.; Альтерио и др., 2009 г.; Пасторек и Пасторекова, 2015; Заманова и др., 2019; Мишра и др., 2020). Экспрессия CA-IX активируется при гипоксии фактором транскрипции, индуцируемым гипоксией-1 (HIF1α), и связана с эффектом Варбурга при онкологической патологии (Ivanov et al., 2001; Supuran, 2008; Pastorek and Pastorekova, 2015; Шабана и Илиес, 2019). На моделях глиобластомы было обнаружено, что CA-IX увеличивает миграцию, подвижность и адгезию клеток в моноцитах (Huang et al. , 2020). В настоящее время соединение, частично селективное в отношении ингибирования CA-IX, обозначенное как SLC-0111, проходит клинические испытания, и вполне вероятно, что оно будет действовать в качестве терапии многих видов рака, включая глиобластому (Boyd et al., 2017; Ilies и Винум, 2019 г.; Шабана и Илиес, 2019 г.; Макдональд и др., 2020). Предполагается также, что эта изоформа участвует в фиброзе сердца в модели лигирования сердца у крыс (Vargas et al., 2016). Интересно, что у людей CA-IX также экспрессируется в атеросклеротических бляшках, где предполагается, что он является маркером некротической ткани (Demandt et al., 2021). Недавние исследования предоставили доказательства того, что CA-IX, вероятно, усугубляет исходы церебральной ишемии (Mishra et al., 2020). CA-XII — еще один среднескоростной (K кат. = 4,2 × 10 5 с –1 ), мембраносвязанный изофермент, сходный по общей структуре с CA-IX, но без протеогликанового домена. Он также димерный, с двумя активными центрами, ориентированными во внеклеточную среду (Ivanov et al. , 1998; Tureci et al., 1998; Whittington et al., 2001). Он обнаруживается во многих гипоксических опухолях, наряду с CA-IX, включая глиомы, гемангиобластомы и менингиомы. Однако, в отличие от СА-IX, СА-XII в высокой степени экспрессируется во многих нормальных тканях, включая эпителий молочной железы и непигментированные цилиарные эпителиальные клетки глаза, в головном мозге (мелкие периферические капилляры), пищеводе, поджелудочной железе, толстой кишке, прямой кишке, почках, простата, яичники, яички, эндометрий, потовые железы. Было показано, что CA-XII играет ключевую роль в электролитном гомеостазе эпителиальных клеток посредством активации протоков Cl 9.0792 – /HCO 3 – обменник AE2 и ассоциация с Na + /HCO 3 – котранспортер kNBC1 (Proescholdt et al., 2005; и др., 2015; Вахид и Слай, 2017; Заманова и др., 2019; Мишра и др., 2020). Мембраносвязанный изофермент СА-XIV обладает внеклеточным каталитическим доменом, одиночной трансмембранной спиралью, коротким внутриклеточным полипептидным сегментом и обладает умеренной каталитической активностью. Он сильно экспрессируется в почках, сетчатке и сердце, а также в головном мозге, скелетных мышцах, печени и легких. Сообщалось, что CA-XIV взаимодействует с переносчиками бикарбонатов и участвует в кислотно-щелочном балансе в мышцах и эритроцитах в ответ на хроническую гипоксию и гиперактивность сердца (Mori et al., 19).99; Супуран, 2008 г.; Мбоге и др., 2018 г.; Заманова и др., 2019; Мишра и др., 2020).
Митохондриальные изоформы СА-ВА и СА-ВБ являются изоферментами со средне-высокой активностью (К кат ВА = 2,9 × 10 5 с –1 , К × кат ВБ1 10 5 s –1 ), важный для глюконеогенеза, липогенеза, уреагенеза и других анаболических путей. Они поставляют HCO 3 – пируваткарбоксилазе во время путей глюконеогенеза и липогенеза и карбамоилфосфатсинтетазе в пути уреагенеза. CA-VA обнаруживается в основном в печени, тогда как CA-VB обнаруживается в скелетных и сердечных мышцах, почках, поджелудочной железе, желудочно-кишечном тракте, головном и спинном мозге (Shah et al. Хотя в обширных исследованиях сообщалось о множественных функциях различных изоформ КА в различных тканях, необходимы дальнейшие исследования по переоценке клеточно-специфической экспрессии/активности КА, особенно в ЦНС и при нарушениях ЦНС. В частности, еще предстоит выяснить влияние КА на цереброваскулярную дисрегуляцию, возникающую при ИИ и БА. Поэтому крайне важно определить роли, специфичные для клеток и изоформ, и разработать селективные по изоформам ингибиторы, которые, вероятно, улучшат специфичность клеток/тканей и уменьшат наблюдаемые побочные эффекты пан-ИАИ, одобренных FDA. Провенси и др., 2019 г.; Заманова и др., 2019; Мишра и др., 2020). Ниже мы обобщаем доступную литературу, в которой подчеркиваются положительные эффекты ингибирования СА на нейроваскулярную дисфункцию при цереброваскулярной патологии, вызванной инсультом, AD, CAA и диабетом. CAI изучались в течение десятилетий (Maren, 1967; Bertini and Luchinat, 1983; Silverman and Lindskog, 1988; Supuran et al. MTZ, например, был признан одним из немногих соединений, обладающих способностью ингибировать высвобождение цитохрома с из митохондрий при окислительном стрессе (Wang et al. Было показано, что ингибирование CA способствует тонусу сосудов головного мозга во время переходных фаз изменения pH, но не в стационарных условиях в артериях крыс (Rasmussen and Boedtkjer, 2018). Нейропротекторные эффекты МТЗ в обоих случаях in vitro и модели ишемического повреждения in vivo , представленные в таблице 1, были впервые исследованы Wang et al. (2009). Воздействие OGD и H 2 O 2 на первичные корковые нейроны привело к некрозу, который был купирован лечением MTZ. Точно так же OGD увеличивал высвобождение цитохрома с и высвобождение фактора, индуцирующего апоптоз (AIF), из митохондрий, а также активность каспазы-3 (Wang et al., 2009), а MTZ ослаблял эти эффекты (Wang et al., 2009). , предполагая, что MTZ предотвращает не только некроз, но и митохондриально-опосредованный апоптоз, индуцированный OGD. В том же исследовании первичные нейроны коры, подвергшиеся воздействию OGD, демонстрировали активацию воспалительных процессов, измеряемую активацией каспазы-1 и высвобождением IL-1β. MTZ ингибировал как высвобождение IL-1β, так и активацию каспазы-1 in vitro (Wang et al. Таблица 1. Ингибирование КА в моделях ИИ. Таблица 2. Ингибирование КА в моделях ГС. Защитные механизмы ингибирования КА в моделях ГС обобщены в таблице 2. Ингибирование КА уменьшало повреждение головного мозга после ВМК у крыс Sprague-Dawley (Guo et al., 2012). Это исследование было сосредоточено на CA-I, который сильно экспрессируется в эритроцитах. При интракаудальной инъекции крови уровни CA-I повышались в ипсилатеральных базальных ганглиях в течение 3 дней после инъекции. Недавно на моделях амилоидоза было протестировано влияние ингибирования КА на функцию митохондриальных и нервно-сосудистых клеток (таблица 3). При БА и ЦАА токсичные агрегаты Aβ накапливаются вокруг и внутри стенок сосудов головного мозга, а также в паренхиматозных бляшках в определенных областях мозга, таких как гиппокамп и кора (Ghiso et al., 2014; Cortes-Canteli and Iadecola, 2020). . Наша лаборатория показала, что олигомеры и протофибриллы Aβ индуцируют высвобождение цитохрома с из митохондрий во многих типах клеток NVU, включая ECs и SMCs, нейроны и глиальные клетки, что приводит к активации каспазы и гибели клеток (Fossati et al. Таблица 3. Ингибирование СА в моделях БА и САА. Интересно, что хотя CAI доказали свою эффективность в предотвращении патологии CAA и AD, активаторы CA также продемонстрировали положительное влияние на сохранение памяти в острых моделях (Ilies et al., 2002; Canto de Souza et al., 2017; Sanku et al. al., 2018; Provensi et al., 2019; Blandina et al., 2020; Schmidt et al., 2020). Эти данные указывают на важную роль ферментов CA в модуляции множества путей при контекстуальном угасании памяти о страхе, нейродегенеративной и нейроваскулярной патологии. Они также подчеркивают необходимость тщательной и специфической модуляции активности фермента CA и дальнейшего понимания эффектов каждой изоформы в различных областях мозга, типах клеток и патологических состояниях. , 2000; Supuran, 2008; Scozzafava et al., 2013 г. Заманова и др., 2019 г.; Мишра и др., 2020). Из-за их участия в этих анаболических путях их изучали на моделях ожирения и диабета 2 типа (De Simone and Supuran, 2007; Supuran, 2012; Scozzafava et al., 2013; Salameh et al., 2019; Mishra et al. ., 2020). Функция CA-V и различие между изоформами CA-VA и CA-VB были исследованы путем анализа фенотипических различий между CA-VA и CA-VB KO, а также моделей мышей с двойным KO, что указывает на их роль в аммиаке. детоксикации (Shah et al., 2013b). Функция CA-V в головном мозге крайне мало изучена, однако сообщалось, что CA-VA экспрессируется как в нейронах, так и в глиальных клетках (Ghandour et al., 2000). Интересно, что эффект CA-V был также исследован в церебральных перицитах (Price et al., 2017). Было замечено, что молчание обеих изоформ защищает от индуцированной высоким уровнем глюкозы гибели клеток и продукции АФК, CA-VA в более значительной степени (Price et al., 2017), что также подтверждается усилением окислительного стресса и апоптоза в моделях Сверхэкспрессия CA-VA (Price et al. , 2017). Ингибиторы карбоангидразы и цереброваскулярная патология
, 2003; Krishnamurthy et al., 2008; Supuran, 2008). ; Винум и др., 2009 г.; Альтерио и др., 2012; Акочак и Илиес, 2014 г.; Маккенна и Супуран, 2014 г.; Супуран и Винум, 2015 г.; Илиес и Винум, 2019 г.; Анджели и др., 2020 г.; Супуран и Капассо, 2020 г.). Сначала они были разработаны как диуретики из-за их функции по реабсорбции натрия и воды в почках (DuBose, 1984; Purkerson and Schwartz, 2007; Supuran, 2018), и в настоящее время используются для лечения глаукомы, поскольку они снижают внутриглазное давление и для профилактики высотной болезни (Bradwell et al., 1992; Минсионе и др., 2007 г.; Ritchie et al., 2012), поскольку они уменьшают легочную вазоконстрикцию, увеличивают CBF, вероятно, контролируя церебральную оксигенацию, и уменьшают отек головного мозга (Mishra et al., 2020). Однако молекулярные механизмы, ответственные за эффекты CAI, включая некоторые из наиболее часто используемых пан-CAI, такие как ATZ и MTZ, многочисленны и все еще находятся в стадии изучения. , 2008). Более 1000 соединений из библиотеки лекарств NINDS были сначала исследованы в изолированных митохондриях печени мышей, а затем подтверждены в стриарных клетках на моделях болезни Гентингтона. MTZ, ингибируя высвобождение цитохрома с из пораженных митохондрий, также приводил к снижению уровня каспазы-9.и активация каспазы-3 (Wang et al., 2008; Fossati et al., 2016; Sekerdag et al., 2018). После этого исследования одобренные FDA CAI, такие как MTZ, ATZ, топирамат и совсем недавно разработанные селективные ингибиторы, не одобренные FDA, начинают применяться в моделях цереброваскулярной патологии, ишемии, CAA и AD, и их положительные эффекты были приписаны , по крайней мере частично, их способностью предотвращать митохондриальную дисфункцию в цереброваскулярных клетках (Wang et al., 2009; Shah et al., 2013a; Fossati et al., 2016; Solesio et al., 2018). В этом исследовании использовались два разных CAI, ATZ и 4-аминометилбензолсульфонамид (АМБ). Выявлено, что только пан-ЦАИ АТЗ, активный в отношении большинства внутриклеточных изоферментов КА, оказывает влияние на механизмы внутриклеточного закисления. На основании наблюдаемых результатов был сделан вывод, что внутриклеточные CA ответственны за изменение скорости внутриклеточного pH и тонуса сосудов в артериях (Rasmussen and Boedtkjer, 2018). В других исследованиях наблюдались сосудорасширяющие способности АТЗ, такие как повышенная продукция NO, приводящая к усилению кровотока в коре головного мозга крыс (Tuettenberg et al., 2001). Вполне вероятно, что продукция NO — не единственный механизм, с помощью которого АТЗ опосредует вазодилатацию (Kiss et al., 19).99). Системное введение АТЗ также широко использовалось в качестве короткого теста для увеличения CBF в исследованиях на людях (Okudaira et al., 1995; Grossmann and Koeberle, 2000; Russell et al., 2008). Интересно, что его стимулирующий эффект на CBF снижен у пациентов с AD или сосудистой деменцией по сравнению со здоровыми людьми (Stoppe et al. , 1995; Pavics et al., 1999). Ингибирование карбоангидразы в моделях ишемического инсульта
, 2009). Это открытие подтверждает гипотезу о том, что CA могут опосредовать как апоптотические, так и воспалительные механизмы. В мышиной модели MCAO у мышей, получавших MTZ (20 мг/кг), был меньший размер инфаркта, улучшенный неврологический показатель и сниженное высвобождение цитохрома с и активация каспазы-3 по сравнению с мышами, не получавшими лечения (Wang et al., 2009). В более позднем исследовании, проведенном на крысах, была определена эффективность АТЗ отдельно, а также в сочетании с наклоном головы вниз, способом физического стимулирования CBF, в модели транзиторной MCAO у крыс. Результаты показали, что ATZ оказывает защитное действие после транзиторной MCAO, значительно уменьшая размер инфаркта (Han et al., 2020). Кроме того, ATZ снижал экспрессию AQP4 по сравнению с крысами, не получавшими лечения, что свидетельствует об уменьшении отека мозга (Han et al., 2020). В крысиной модели постоянного MCAO ATZ использовали в сравнении с селективными ингибиторами CA-VII, CA-IX и CA-XII, чтобы определить, вовлечены ли эти изоформы в патологию. Интересно, что pan-CAI ATZ не улучшал неврологический показатель через 24 часа после окклюзии, однако CAI с селективностью в отношении CA-VII, а также соединение, более селективное в отношении мембранных изоформ CA-IX и CA-XII, улучшали неврологический дефицит. наблюдали у окклюзированных необработанных крыс. В группе, получавшей мощный и селективный ингибитор CA-IX/CA XII, 6-(бензилокси)бензо[d]тиазол-2-сульфонамид (ББТ), размер инфаркта был значительно уменьшен, в то время как в CA не наблюдалось уменьшения -VII группа ингибиторов. Однако у крыс, получавших 50 мг/кг ATZ, также наблюдалось уменьшение объема инфаркта. Это исследование предполагает, что CA-IX и, возможно, CA-XII могут усугублять потерю нейронов наряду с неврологическим и сосудистым дефицитом после ишемического инсульта (Di Cesare Mannelli et al., 2016). Совсем недавно в другом исследовании оценивались различные СА-селективные ингибиторы как в in vitro и in vivo модели ИС (Dettori et al., 2021). В этом исследовании проанализировано новое поколение CAI, которые являются липофильными и селективными в отношении ассоциированных с гипоксией (CA-IX и CA-XII) и цитозольных (CA-II и CA-I) изоформ. Важно отметить, что в этом исследовании в качестве эталонного соединения использовали АТЗ в гораздо более низкой дозе (4,4 мг/кг). In vitro , липофильные CAI и ATZ защищали от аноксической деполяризации, вызванной OGD, предоставляя механистическую информацию. В естественных условиях , CAI уменьшали объем инфаркта, неврологический дефицит, повреждение нейронов и активацию микроглии (Dettori et al., 2021). В этом исследовании также были измерены уровни TNF-α и IL-10 в плазме, показывающие различия между группами с плацебо-операцией и группами с окклюзией без какого-либо влияния на обработанные группы, вероятно, из-за короткого временного окна лечения (24 часа). Как предполагалось в ходе обсуждения, возможно, что при более длительном лечении ингибирование КА может опосредовать воспалительные пути (Bulli et al., 2021; Dettori et al., 2021). Действительно, СА также связаны с воспалительными патологиями, такими как ревматоидный артрит и метастазирование рака (Supuran, 2008; Alver et al. , 2011; Liu et al., 2012). В целом, как МТЗ, так и АТЗ обладают защитным действием на моделях ИИ, однако разработка ингибиторов СА-IX и, возможно, ингибиторов СА-XII может быть полезной для лечения как неврологических, так и сосудистых расстройств после ИИ. Защитные механизмы, наблюдаемые после ингибирования СА в моделях ИИ, суммированные в таблице 2, по-видимому, превосходят соединения, селективные в отношении мембраносвязанных изоформ СА-IX и СА-XII (Bulli et al., 2021). Ингибирование карбоангидразы в моделях геморрагического инсульта
Кроме того, после внутрихвостовой инъекции CA-I было обнаружено повышенное содержание воды в мозге, активация микроглии и гибель нейронов, что позволяет предположить, что экспрессия CA-I может способствовать отеку мозга и нейровоспалению. У крыс, получавших ATZ, наблюдалось уменьшение перигематомного отека, а также накопление натрия вместе со снижением нейронального дефицита (Guo et al., 2012). Однако клеточные механизмы, ответственные за влияние CA-I на целостность ГЭБ, требуют дальнейшего изучения. В последнее время научное сообщество начинает выдвигать гипотезу о том, что CA-I как в сетчатке, так и в мозге способствует проницаемости сосудов, предполагая, что специфические ингибиторы CA-I могут быть полезными для минимизации дисфункции NVU и, следовательно, нейровоспаления вокруг барьера (Gao и др., 2007). Кроме того, в мышиной модели SAH у мышей, получавших пан-CAI, MTZ, наблюдалось снижение активации каспазы-3 и апоптоза в гиппокампе и коре головного мозга по сравнению с группой, не получавшей лечения, с улучшением неврологического дефицита (Li et al. , 2016). ). В первичных корковых нейронах MTZ снижал гемоглобин и индуцированную кровью гибель клеток наряду с продукцией АФК (Li et al., 2016). Топирамат, другой CAI, активный в отношении нескольких ферментов CA, включая CA-V, уменьшал повреждение SAH у кроликов (Seçkin et al., 2009).). Ингибирование карбоангидразы в моделях болезни Альцгеймера и церебральной амилоидной ангиопатии
, 2010, 2012a). ,b, 2016; Пароди-Руллан и др., 2020). Во всех этих типах нервно-сосудистых клеток MTZ уменьшал митохондриально-опосредованную гибель клеток, запускаемую Aβ (Fossati et al., 2010, 2016). Кроме того, MTZ ослаблял каспазу-3 и каспазу-9.активация в ЭК, нейрональных и глиальных клетках in vitro , а также активация каспазы-3 in vivo (Fossati et al., 2016; Solesio et al., 2018). В исследовании in vivo Aβ вводили в гиппокамп мышей дикого типа в присутствии или в отсутствие предшествующей внутрибрюшинной инъекции MTZ. Интересно, что лечение MTZ ослабляло активацию каспазы-3 в микроглии и увеличивало количество NeuN-позитивных нейронов в гиппокампе, указывая на то, что лечение MTZ оказывало защитный эффект против нейродегенерации после инъекции Aβ (Fossati et al., 2016). Чтобы выяснить конкретные механизмы, ответственные за эти защитные эффекты, наша группа показала, что Aβ-индуцированная митохондриальная дисфункция, деполяризация митохондриальной мембраны, высвобождение цитохрома C и H 2 O 2 ослаблялись в нейронах и ЭК не только MTZ, но и ATZ, который был эффективен при более низких концентрациях (Solesio et al. , 2018). Более того, как ATZ, так и MTZ ингибировали активацию каспазы-9, вызванную Aβ в цереброваскулярных EC, и возникающий в результате апоптоз (Solesio et al., 2018). Также было замечено, что MTZ увеличивает активацию фактора 2, связанного с ядерным фактором (Nrf2), в моделях высотной болезни и, что более интересно, в клетках нейробластомы человека и первичных корковых нейронах, зараженных Aβ 9.1650 in vitro (Lu et al., 2020; Sotolongo et al., 2020). Активация Nrf2 с помощью MTZ повышала активность антиоксидантных ферментов, таких как супероксиддисмутаза-1 и гем-оксигеназа-1, что указывает на потенциальные побочные эффекты MTZ (Sotolongo et al., 2020). Эти исследования подтверждают гипотезу о том, что ингибирование СА является защитным для нескольких типов клеток NVU в моделях амилоидоза, и подчеркивают необходимость тестирования этих и/или специфических для изоформ соединений в клинических испытаниях при БА и САА. Новые исследования, проводимые в настоящее время в нашей лаборатории, также подтверждают положительное влияние CAI на когнитивные функции в мышиных моделях амилоидоза (Angiulli et al. , 2018). В настоящее время проводятся дополнительные исследования для изучения хронического лечения пан-CAI на нескольких моделях CAA и AD, параллельно с оценкой специфических ингибиторов изоформ, для дальнейшего выяснения роли различных изоформ CA в патологии AD и CAA. Ингибирование карбоангидразы в моделях вызванной диабетом патологии сосудов головного мозга различные модели диабета (Price et al., 2012, 2017; Shah et al., 2013a), который способствует развитию болезни Альцгеймера и инсульта (Kuźma et al., 2018; Cortes-Canteli and Iadecola, 2020; Freitas-Andrade et al. ., 2020; Злокович и др., 2020). В модели мышей с диабетом CD-1, индуцированным стрептозотоцином, сообщалось, что лечение топираматом в течение 3 недель повышало уровень глутатиона и уменьшало окислительный стресс в головном мозге. Также было замечено, что индуцированная диабетом потеря церебральных перицитов ослаблялась топираматом (Price et al., 2012). Та же группа подтвердила эти механизмы в мозге мышей CA-V с двойным нокаутом, где наблюдались повышенные уровни глутатиона и сниженный окислительный стресс по сравнению с диким типом (Price et al., 2012). Обработанные гипергликемией цереброваскулярные перициты увеличивали продукцию АФК, а также скорость митохондриального дыхания.
Ингибирование CA (особенно CA-V) топираматом снижало продукцию АФК (Shah et al., 2013a). Хотя топирамат обладает высокой ингибирующей активностью в отношении изоформ CA-V, он также обладает сродством к другим изоформам, а также множественными нецелевыми эффектами в головном мозге, как показали исследования, в которых анализировались функции топирамата в отношении рецепторов ГАМК и глутамата (Johnson , 2005; Мао и др., 2015). В идеале следует разработать более специфические ингибиторы для дальнейшего изучения функций CA-VA/B. Та же группа использовала плазмиду для сверхэкспрессии CA-VA в перицитах головного мозга, подвергшихся воздействию высокого уровня глюкозы, чтобы подтвердить предполагаемые молекулярные механизмы, связанные с CA-VA, такие как продукция АФК и гибель клеток, отметив, что топирамат защищает от этих механизмов (Patrick et al. ., 2015). В качестве доказательства концепции та же группа генетически сбивала CA-VA и CA-VB в церебральных перицитах (Price et al., 2017). Нокдаун CA-VA и CA-VB улучшал жизнеспособность клеток по сравнению с контролем при воздействии гипергликемии, хотя специфический нокдаун CA-VA был даже более эффективным, чем нокдаун CA-VB, в снижении продукции АФК и апоптоза в условиях гипергликемии. (Прайс и др., 2017). Эти исследования выявили потенциальные различия между функциями CA-VA и CA-VB в перицитах головного мозга, которые требуют дальнейшего изучения. В мышиной модели диабета, индуцированного стрептозотоцином, цереброваскулярную патологию также охарактеризовали с помощью электронной микроскопии. Разрушение ГЭБ и его дисфункция уменьшались при лечении топираматом in vivo (Salameh et al., 2016). Совсем недавно исследование с использованием мышиной модели диабета с высоким содержанием жиров было сосредоточено на нарушении гиппокампального ГЭБ. Белки плотного соединения BBB, такие как ZO-1 и клаудин-12, снижались при диете с высоким содержанием жиров, в то время как лечение топираматом увеличивало их экспрессию наряду с ослаблением окислительного стресса (Salameh et al., 2019).
Заключение
НВУ является важной функциональной структурой ЦНС, и ее недостаточность участвует в развитии БА, сосудистой деменции и усугубляет исходы инсульта. Множественные патологические и клеточные механизмы, приводящие к патологии NVU, нуждаются в прояснении, а новые терапевтические стратегии должны быть дополнительно исследованы и разработаны. По мере совершенствования научных методов увеличивается способность понимать функции этого блока. Недавние исследования подтверждают, что ингибирование CA является защитным для NVU, и роль этих ферментов требует дальнейшего изучения. В таблице 4 подчеркнуты изоформы CA, которые обсуждались на протяжении всего обзора, их клеточная локализация, их связь с конкретными неврологическими расстройствами и их экспрессия в сосудисто-нервных клетках. Выяснение того, как клетки NVU взаимодействуют друг с другом, а также роли CA в каждом типе клеток, имеет решающее значение для нацеливания на цереброваскулярную патологию при ИИ, AD, а также других нейрогенеративных заболеваниях. Пан-CAI MTZ и ATZ, а также топирамат продемонстрировали защитный эффект на моделях инсульта, цереброваскулярной патологии, диабета II типа и болезни Альцгеймера, что обобщено на рисунке 2. Хотя эти соединения одобрены FDA, они облегчают переход к клиническим испытаниям. , эти CAI не являются специфическими препаратами. CAI, специфичные для изоформ, вызывают все больший интерес, поскольку у человека было идентифицировано 15 изоформ с различными функциями и локализациями. Необходимы дальнейшие доклинические и клинические исследования для оценки эффективности ингибирования КА при БА, а также при ИИ. Для подтверждения роли специфических изоформ CA в качестве фармакологических мишеней были бы полезны генетические исследования, такие как изоформа KO и нокаут в различных типах клеток NVU. В целом, обсуждавшиеся выше исследования предоставляют доказательства того, что КА являются важными потенциальными медиаторами и мишенями в нейроваскулярной патологии при БА, инсульте и связанных с ними цереброваскулярных нарушениях.
Таблица 4. Изоформы КА при ЦНС и цереброваскулярной патологии.
Рисунок 2. AD и инсульт имеют общие факторы риска и патологические механизмы. Этот рисунок направлен на то, чтобы подчеркнуть множественные CAI, а также указать на конкретные изоформы CA, которые, как наблюдалось, модулируют цереброваскулярную патологию в моделях AD или инсульта. Возраст, сердечно-сосудистые факторы риска и гипоперфузия — все это общие факторы риска развития БА и инсульта. И БА, и инсульт демонстрируют дисфункцию NVU, сопровождающуюся многими общими молекулярными механизмами, такими как митохондриальная дисфункция, гибель сосудов и нейронов, нейровоспаление и дисфункция ГЭБ. Являются ли эти механизмы причинами или следствиями нейроваскулярной дисфункции, до сих пор остается предметом горячих дискуссий. Несмотря на различия в поздних патологических проявлениях двух заболеваний, инсульт действительно увеличивает риск деменции, что позволяет предположить, что воздействие на их общие факторы риска и молекулярные механизмы может в конечном итоге смягчить развитие цереброваскулярной патологии при обоих заболеваниях, защищая здоровье мозга и гомеостаз ЦНС. АТЗ, ацетазоламид; МТЗ, метазоламид; ГЭБ, гематоэнцефалический барьер; NVU, сосудисто-нервная единица; ЦНС, центральная нервная система; CA, карбоангидраза; AD, болезнь Альцгеймера; CAIs, ингибиторы карбоангидразы. Эта фигура была создана с помощью BioRender.com.
Вклад авторов
NL и SF разработали обзор и разработали его концепцию. NL написал черновой вариант обзора и провел поиск литературы. SF критически пересмотрела, отредактировала рукопись, предоставила соответствующие идеи, провела дополнительный поиск литературы и получила финансирование. EC и MI пересмотрели рукопись и провели дополнительный поиск литературы. Все авторы внесли свой вклад в статью и одобрили представленную версию.
Финансирование
Эта работа была поддержана грантами NIH R01NS104127 и R01AG062572, Программой наград Мемориального фонда Эдварда Н. и Деллы Л. Том в области исследования лекарств от болезни Альцгеймера, Ассоциацией Альцгеймера (AARG), Департаментом здравоохранения Пенсильвании. на грант болезни Альцгеймера (PA Cure), присужденный SF, благотворительным фондом Карен Тоффлер и Лемолевским центром комплексных лимфатических исследований.
Конфликт интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Примечания издателя
Все претензии, изложенные в этой статье, принадлежат исключительно авторам и не обязательно представляют претензии их дочерних организаций или издателя, редакторов и рецензентов. Любой продукт, который может быть оценен в этой статье, или претензии, которые могут быть сделаны его производителем, не гарантируются и не поддерживаются издателем.
Благодарности
Мы благодарим Эшли Кэри за концептуализацию и рисование рисунка 1 обзора. Мы также признаем, что Рисунок 2 был создан с использованием BioRender.com.
Ссылки
Akocak, S., Alam, M.R., Shabana, A.M., Sanku, R.K., Vullo, D., Thompson, H., et al. (2016). Пегилированные бис-сульфонамидные ингибиторы карбоангидразы могут эффективно контролировать рост нескольких карцином, экспрессирующих карбоангидразу IX. J. Med. хим. 59, 5077–5088. doi: 10.1021/acs.jmedchem.6b00492
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Акочак С. и Илиес Массачусетс (2014). «Первичные сульфонамидные ингибиторы карбоангидразы нового поколения» в Targeting Carbon Anhydrases. (Eds) CT Supuran. и C. Cappasso. (Лондон: Future Science Ltd). 34–51. doi: 10.4155/fseb2013.13.22
Полный текст CrossRef | Google Scholar
Алдеа Р., Веллер Р. О., Уилкок Д. М., Караре Р. О. и Ричардсон Г. (2019). Цереброваскулярные гладкомышечные клетки как факторы интрамурального периартериального дренирования головного мозга. Фронт. Стареющие нейроски. 11:1.
Google Scholar
Аллури Х., Стэгг Х.В., Уилсон Р.Л., Клейтон Р.П., Савант Д.А., Конеру М. и др. (2014). Взаимосвязь активных форм кислорода и каспазы-3 в опосредовании гиперпроницаемости эндотелиальных клеток гематоэнцефалического барьера после кислородно-глюкозной депривации и реоксигенации. Микроциркуляция 21, 187–195.
Google Scholar
Альтерио, В., Ди Фиоре, А., Дамброзио, К., Супуран, К. Т., и Де Симоне, Г. (2012). Множественные способы связывания ингибиторов карбоангидразы: как разработать специфические препараты, нацеленные на 15 различных изоформ? хим. Ред. 112, 4421–4468. doi: 10.1021/cr200176r
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Альтерио В., Хилво М., Ди Фиоре А., Супуран С. Т., Пан П., Парккила С. и др. (2009). Кристаллическая структура каталитического домена ассоциированной с опухолью карбоангидразы IX человека. Проц. Натл. акад. науч. США 106, 16233–16238. doi: 10.1073/pnas.01106
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Алвер А., Шентюрк А., Чакирбай Х., Ментеше А., Гёкмен Ф., Кеха Э. Э. и др. (2011). Аутоантитела к карбоангидразе II и окислительный стресс при ревматоидном артрите. клин. Биохим. 44, 1385–1389. doi: 10.1016/j.clinbiochem.2011.09.014
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Андреоне, Б. Дж., Лакост, Б., и Гу, К. (2015). Нейрональные и сосудистые взаимодействия. год. Преподобный Нейроски. 38, 25–46. doi: 10.1146/annurev-neuro-071714-033835
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Андринг Дж. Т., Фууч М., Акочак С., Анджели А., Супуран С. Т., Илиес М. А. и др. (2020). Структурные основы наномолярного ингибирования ассоциированной с опухолью карбоангидразы IX: рентгеноструктурное исследование и исследование ингибирования липофильных ингибиторов с ацетазоламидным остовом. J. Med. хим. 63, 13064–13075. doi: 10.1021/acs.jmedchem.0c01390
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ангели А., Карта Ф., Ночентини А., Винум Дж. Ю., Залюбовскис Р., Акдемир А. и др. (2020). Ингибиторы карбоангидразы, воздействующие на метаболизм и микроокружение опухоли. Метаболиты 10:412. doi: 10.3390/metabo10100412
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Анджели А., Вайано Ф., Мари Ф., Бертол Э. и Супуран К.Т. (2017). Психоактивные вещества, принадлежащие к классу амфетаминов, сильно активируют изоформы VA, VB, VII и XII карбоангидразы мозга. Дж. Ингибитор ферментов. Мед. хим. 32, 1253–1259. doi: 10.1080/14756366.2017.1375485
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ангиулли Ф., Солезио М. Э., Дебуре Л., Сехудо Дж. Р., Вишневски Т. и Фоссати С. (2018). P3-464: ингибиторы карбоангидразы улучшают нейроваскулярную дисфункцию в мышиной модели церебральной амилоидной ангиопатии. Альцгеймер Демент. 14, 1296–1296. doi: 10.1016/j.jalz.2018.06.1828
CrossRef Полный текст | Академия Google
Аспатвар, А., Толванен, М.Е., Ортутай, К., и Парккила, С. (2014). Белки, родственные карбоангидразе: молекулярная биология и эволюция. Субъячейка. Биохим. 75, 135–156. doi: 10.1007/978-94-007-7359-2_8
Полный текст CrossRef | Google Scholar
Остин С.А., Сантанам А.В., Хинтон Д.Дж., Чой Д.С. и Катусик З.С. (2013). Эндотелиальный дефицит оксида азота способствует патологии болезни Альцгеймера. Дж. Нейрохим. 127, 691–700. doi: 10.1111/jnc.12334
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Бадимон А. , Страсбургер Х. Дж., Аята П., Чен X., Наир А., Икегами А. и др. (2020). Отрицательная обратная связь контроля активности нейронов с помощью микроглии. Природа 586, 417–423. doi: 10.1038/s41586-020-2777-8
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Бейнс, Дж. С., и Шоу, Калифорния (1997). Нейродегенеративные расстройства у людей: роль глутатиона в опосредованной окислительным стрессом гибели нейронов. Brain Res.Rev. 25, 335–358. doi: 10.1016/S0165-0173(97)00045-3
Полный текст CrossRef | Google Scholar
Бертини И. и Лучинат К. (1983). Кобальт(II) как зонд структуры и функции карбоангидразы. Счета хим. Рез. 16, 272–279. doi: 10.1021/ar00092a002
CrossRef Полный текст | Google Scholar
Бирнаски Дж., Корбетт Д., Пилинг Дж., Уэллс Дж. и Лей Х. (2001). Серийное МР-исследование изменений мозгового кровотока и развития повреждений после эндотелин-1-индуцированной ишемии у крыс. Магн. Резон Мед. 46, 827–830. doi: 10.1002/mrm.1263
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Бландина П., Провенси Г., Пассани М. Б., Капассо К. и Супуран К. Т. (2020). Модуляция карбоангидразы эмоциональной памяти. Последствия для лечения когнитивных расстройств. J. Ингибитор ферментов. Мед. хим. 35, 1206–1214. doi: 10.1080/14756366.2020.1766455
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Боспфлюг, Э. Л., и Илифф, Дж. Дж. (2018). Возникающая связь между обменом интерстициальной жидкости и спинномозговой жидкости, бета-амилоидом и сном. биол. Психиатрия 83, 328–336. doi: 10.1016/j.biopsych.2017.11.031
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Боланд Б., Ю У. Х., Корти О., Мольеро Б., Энрикес А., Безар Э. и др. (2018). Способствует выведению нейротоксических белков при нейродегенеративных заболеваниях старения. Нац. Преподобный Друг Дисков. 17, 660–688. doi: 10.1038/nrd.2018.109
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Бориак-Шодин, П. А., Хек, Р. В., Лайпис, П. Дж., Сильверман, Д. Н., и Кристиансон, Д. В. (1995). Определение структуры мышиной митохондриальной карбоангидразы V с разрешением 2,45 A: последствия для каталитического переноса протона и дизайна ингибитора. Проц. Натл. акад. науч. США 92, 10949–10953. doi: 10.1073/pnas.92.24.10949
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Boyd, N.H., Walker, K., Fried, J., Hackney, J.R., Mcdonald, P.C., Benavides, G.A., et al. (2017). Добавление ингибитора карбоангидразы 9 SLC-0111 к лечению темозоломидом задерживает рост глиобластомы in vivo. JCI Insight 2:e. doi: 10.1172/jci.insight.
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Брэдвелл А. Р., Райт А. Д., Винтерборн М. и Имрей К. (1992). Ацетазоламид и высотные болезни. Междунар. Дж. Спорт Мед. 13, С63–С64. doi: 10.1055/s-2007-1024597
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Браун М. и Илифф Дж. Дж. (2020). Влияние нейроваскулярной, гематоэнцефалической барьерной и лимфатической дисфункции при нейродегенеративных и метаболических заболеваниях. Междунар. Преподобный Нейробиол. 154, 413–436. doi: 10.1016/bs.irn.2020.02.006
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Булли И., Деттори И., Коппи Э., Черчи Ф., Вентурини М., Ди Чезаре Маннелли Л. и др. (2021). Роль карбоангидразы в церебральной ишемии и ингибиторов карбоангидразы как предполагаемых защитных агентов. Междунар. Дж. Мол. науч. 22:5029. doi: 10.3390/ijms22095029
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Канепа Э. и Фоссати С. (2020). Влияние тау на нейроваскулярную патологию при болезни Альцгеймера. Фронт. Нейрол. 11:573324. doi: 10.3389/fneur.2020. 573324
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Canto de Souza, L., Provensi, G., Vullo, D., Carta, F., Scozzafava, A., et al. (2017). Активация карбоангидразы улучшает память распознавания объектов у мышей за счет фосфорилирования киназы, регулируемой внеклеточным сигналом, в коре и гиппокампе. Нейрофармакология 118, 148–156. doi: 10.1016/j.neuropharm.2017.03.009
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Караре Р. О., Алдеа Р., Агарвал Н., Баскай Б. Дж., Бекман И., Бош Д. и др. (2020). Клиренс интерстициальной жидкости (ISF) и CSF (CLIC) — часть группы сосудистых профессиональных интересов (PIA): цереброваскулярные заболевания и нарушение выведения амилоида-β из головного мозга и сетчатки с возрастом и болезнь Альцгеймера — возможности для терапии. Демент Альцгеймера. 12:e12053. doi: 10.1002/dad2.12053
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Караре, Р. О., Бернардес-Сильва, М., Ньюман, Т. А., Пейдж, А. М., Николл, Дж. А., Перри, В. Х., и др. (2008). Растворенные вещества, но не клетки, дренируются из паренхимы головного мозга вдоль базальных мембран капилляров и артерий: значение для церебральной амилоидной ангиопатии и нейроиммунологии. Невропатология. заявл. Нейробиол. 34, 131–144. дои: 10.1111/j.1365-2990.2007.00926.x
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Карсель-Маркес Дж., Каллелл Н., Муино Э., Гальего-Фабрега К., Льедос М., Ибаньес Л. и др. (2021). Причинное влияние уровней ММР-1 (матриксная металлопротеиназа-1), ММР-8 и ММР-12 на ишемический инсульт: менделевское рандомизированное исследование. Ход 52, e316–e320. doi: 10.1161/STROKEAHA.120.033041
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Castillo-Carranza, D.L., Nilson, A.N., Van Skike, C.E., Jahrling, J.B., Patel, K., Garach, P., et al. (2017). Церебральное микрососудистое накопление олигомеров тау при болезни Альцгеймера и родственных тауопатиях. Старение Дис. 8, 257–266. doi: 10.14336/AD.2017.0112
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Чеслер, М. (2003). Регуляция и модуляция pH в головном мозге. Физиол. 83, 1183–1221. doi: 10.1152/physrev.00010.2003
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Коррайни П., Хендерсон В. В., Ординг А. Г., Педерсен Л., Хорват-Пухо Э. и Соренсен Х. Т. (2017). Долгосрочный риск деменции среди выживших после ишемического или геморрагического инсульта. Инсульт 48, 180–186. doi: 10.1161/STROKEAHA.116.015242
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Кортес-Кантели, М., и Ядекола, К. (2020). Болезнь Альцгеймера и сосудистое старение: фокус-семинар JACC. Дж. Ам. Сб. Кардиол. 75, 942–951. doi: 10.1016/j.jacc.2019.10.062
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Крус Эрнандес Дж. К., Брако О., Керсберген С. Дж., Мьюз В., Хафт-Джавахериан М., Берг М. и др. (2019). Адгезия нейтрофилов в капиллярах головного мозга снижает кровоток в коре головного мозга и ухудшает функцию памяти в моделях мышей с болезнью Альцгеймера. Нац. Неврологи. 22, 413–420. doi: 10.1038/s41593-018-0329-4
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Да Мескита С., Луво А., Ваккари А., Смирнов И., Корнелисон Р. К. и Кингсмор К. М. (2018). Функциональные аспекты менингеальных лимфатических сосудов при старении и болезни Альцгеймера. Природа 560, 185–191. doi: 10.1038/s41586-018-0368-8
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Дэй, Р. Дж., Мейсон, М. Дж., Томас, К., Пун, В. В., и Рон, Т. Т. (2015). Тау-белок, расщепленный каспазой, совместно локализуется с ранними маркерами клубков в мозге человека с сосудистой деменцией. PLoS One 10:e0132637. doi: 10.1371/journal.pone.0132637
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
де Брюйн, Р. Ф., и Икрам, М. А. (2014). Сердечно-сосудистые факторы риска и будущий риск болезни Альцгеймера. БМС Мед. 12:130. doi: 10.1186/s12916-014-0130-5
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Де Симон Г. и Супуран К. Т. (2007). Ингибиторы карбоангидразы против ожирения. Курс. Верхний. Мед. хим. 7, 879–884. doi: 10.2174/156802607780636762
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Дежакайсая, Х., Кван, П., и Джонс, Северная Каролина (2021). Участие астроцитов и глутамата в патогенезе эпилепсии при болезни Альцгеймера. Эпилепсия 62, 1485–1493. doi: 10.1111/epi.16918
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Демандт, Дж. А. Ф., Дюбуа, Л. Дж., Ван Куйк, К., Затовичова, М., Джин, Х., Парккила, С., и др. (2021). Сенсор гипоксии карбоангидраза IX влияет на метаболизм макрофагов, но не является подходящим биомаркером сердечно-сосудистых заболеваний человека. наук. Респ. 11:425. doi: 10.1038/s41598-020-79978-5
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Деттори И., Фуско И., Булли И., Гавиано Л., Коппи Э., Черчи Ф. и др. (2021). Защитные эффекты ингибирования карбоангидразы при ишемии головного мозга. J. Ингибитор ферментов. Мед. хим. 36, 964–976. doi: 10.1080/14756366.2021.1 5
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Di Cesare Mannelli, L., Micheli, L., Carta, F., Cozzi, A., Ghelardini, C., and Supuran, C. T. (2016). Ингибирование карбоангидразы для лечения церебральной ишемии: оценка in vivo ингибиторов сульфонамида и кумарина. Дж. Ингибитор ферментов. Мед. хим. 31, 894–899. doi: 10.3109/14756366.2015.1113407
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Драгичи Б., Вулло Д., Акочак С., Уокер Э. А., Супуран С. Т. и Илиес М. А. (2014). Этиленбисимидазолы являются сильнодействующими и селективными активаторами изоферментов VA и VII карбоангидразы с потенциальным ноотропным эффектом. Хим. коммун. 50, 5980–5983. doi: 10.1039/C4CC02346C
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
DuBose, TD Jr. (1984). Карбоангидразозависимый транспорт бикарбонатов в почках. Энн. Н. Я. акад. науч. 429, 528–537. doi: 10.1111/j.1749-6632.1984.tb12382.x
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Эльдахшан В., Фаган С. К. и Эргул А. (2019). Воспаление в сосудисто-нервной единице: внимание к микроглии при инсульте и восстановлении. Фармакол. Рез. 147:104349. doi: 10.1016/j.phrs.2019.104349
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Энгельхардт С., Аль-Ахмад А. Дж., Гассманн М. и Огуншола О. О. (2014). Гипоксия избирательно разрушает комплексы плотных соединений эндотелия микрососудов головного мозга через механизм, зависимый от фактора-1, индуцируемого гипоксией (HIF-1). Дж. Сотовый. Физиол. 229, 1096–1105. doi: 10.1002/jcp.24544
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Фарако Г. , Фоссати С., Бьянки М. Э., Патроне М., Педрацци М., Спараторе Б. и др. (2007). Высокоподвижный белок group box 1 высвобождается нервными клетками при различных стрессах и усугубляет ишемическую нейродегенерацию in vitro и in vivo. Дж. Нейрохим. 103, 590–603. doi: 10.1111/j.1471-4159.2007.04788.x
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ферреро Дж., Уильямс Л., Стелла Х., Лейтерманн К., Микульскис А., О’Горман Дж. и др. (2016). Первое двойное слепое плацебо-контролируемое исследование эскалации однократной дозы адуканумаба (BIIB037) на людях при болезни Альцгеймера легкой и средней степени тяжести. Альцгеймер Демент. 2, 169–176. doi: 10.1016/j.trci.2016.06.002
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Фоссати, С., Кэм, Дж., Мейерсон, Дж., Межерихер, Э., Ромеро, И.А., Куро, П.О., и соавт. (2010). Дифференциальная активация путей апоптоза митохондрий васкулотропными вариантами бета-амилоида в клетках, составляющих стенки сосудов головного мозга. FASEB J. 24, 229–241. doi: 10.1096/fj.09-139584
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Фоссати С., Гизо Дж. и Ростаньо А. (2012a). Изучение путей апоптоза, опосредованных каспазой, индуцированных бета-амилоидом в эндотелиальных клетках микрососудов головного мозга. Нейродегенер. Дис. 10, 324–328. doi: 10.1159/000332821
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Фоссати С., Гизо Дж. и Ростаньо А. (2012b). Рецепторы смерти TRAIL DR4 и DR5 опосредуют апоптоз эндотелиальных клеток микрососудов головного мозга, индуцированный олигомерным Aβ болезни Альцгеймера. Раствор клеточной смерти. 3, е321. doi: 10.1038/cddis.2012.55
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Фоссати С., Джаннони П., Солезио М. Э., Коклин С. Л., Кабрера Э., Гизо Дж. и др. (2016). Ингибитор карбоангидразы метазоламид предотвращает индуцированную бета-амилоидом митохондриальную дисфункцию и активацию каспазы, защищая нейроны и глиальные клетки in vitro и в мозге мыши. Нейробиол. Дис. 86, 29–40. doi: 10.1016/j.nbd.2015.11.006
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Фрейтас-Андраде, М., Раман-Наир, Дж., и Лакост, Б. (2020). Структурное и функциональное ремоделирование сосудов головного мозга после инсульта. Фронт. Физиол. 11:948. doi: 10.3389/fphys.2020.00948
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Fujimura, M., Gasche, Y., Morita-Fujimura, Y., Massengale, J., Kawase, M. и Chan, P. H. (1999). Раннее появление активированной матриксной металлопротеиназы-9 и нарушение гематоэнцефалического барьера у мышей после очаговой церебральной ишемии и реперфузии. Мозг Res. 842, 92–100. doi: 10.1016/S0006-8993(99)01843-0
Полный текст CrossRef | Google Scholar
Гао Б. Б., Клермон А., Рук С., Фонда С. Дж., Шринивасан В. Дж., Войтковски М. и др. (2007). Внеклеточная карбоангидраза опосредует геморрагическую проницаемость сетчатки и сосудов головного мозга посредством активации прекалликреина. Нац. Мед. 13, 181–188. doi: 10.1038/nm1534
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Жервуа П. и Ламбрихтс И. (2019). Новая роль триггерного рецептора, экспрессируемого на миелоидных клетках 2, в качестве мишени для иммуномодуляции при ишемическом инсульте. Фронт. Иммунол. 10:1668. doi: 10.3389/fimmu.2019.01668
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Гандур М.С., Лэнгли О.К., Чжу С.Л., Вахид А. и Слай В.С. (1992). Карбоангидраза IV на эндотелиальных клетках капилляров головного мозга: маркер, связанный с гематоэнцефалическим барьером. Проц. Натл. акад. науч. США 89, 6823–6827. doi: 10.1073/pnas.89.15.6823
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Гандур М.С., Парккила А.К., Парккила С., Вахид А. и Слай В.С. (2000). Митохондриальная карбоангидраза в нервной системе: экспрессия в нейрональных и глиальных клетках. Дж. Нейрохим. 75, 2212–2220. дои: 10.1046/j.1471-4159.2000.0752212.x
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Гизо, Дж., Фоссати, С., и Ростаньо, А. (2014). Амилоидоз, связанный с церебральной амилоидной ангиопатией: клеточные сигнальные пути, возникающие в церебральных эндотелиальных клетках. J. Alzheimers Dis. 42, С167–С176. doi: 10.3233/JAD-140027
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Жируар Х. и Ядекола К. (2006). Нейроваскулярная связь в нормальном мозге и при гипертонии, инсульте и болезни Альцгеймера. Дж. Заявл. Физиол. 100, 328–335. doi: 10.1152/japplphysiol.00966.2005
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Гонул Э., Дуз Б., Кахраман С., Каяли Х., Кубар А. и Тимуркайнак Э. (2002). Ранний ответ перицитов на гипоксию головного мозга у кошек: ультраструктурное исследование. Микроваск. Рез. 64, 116–119. doi: 10.1006/mvre.2002.2413
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Гроссманн В. М. и Коберле Б. (2000). Дозозависимая зависимость ацетазоламида от мозгового кровотока у здоровых людей. Церебровасц. Дис. 10, 65–69. doi: 10.1159/000016027
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Го Ф., Хуа Ю., Ван Дж., Кип Р. Ф. и Си Г. (2012). Ингибирование карбоангидразы уменьшает повреждение головного мозга после внутримозгового кровоизлияния. Пер. Инсульт Рез. 3, 130–137. doi: 10.1007/s12975-011-0106-0
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Го, Дж., Краузе, Д. Н., Хорн, Дж., Вайс, Дж. Х., Ли, X., и Даклз, С. П. (2010). Опосредованная эстрогеновыми рецепторами защита жизнеспособности эндотелиальных клеток головного мозга и функции митохондрий после ишемического инсульта in vitro. Дж. Цереб. Кровоток Метаб. 30, 545–554. doi: 10.1038/jcbfm.2009.226
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Гюзель О., Инноченти А., Скоццафава А. , Салман А. и Супуран К. Т. (2009). Ингибиторы карбоангидразы. Фенацетил-, пиридилацетил- и тиенилацетилзамещенные ароматические сульфонамиды действуют как сильнодействующие и селективные ингибиторы изоформы VII. Биоорг. Мед. хим. лат. 19, 3170–3173. doi: 10.1016/j.bmcl.2009.04.123
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Хаапасало, Дж., Нордфорс, К., Хаапасало, Х., и Парккила, С. (2020). Экспрессия карбоангидраз II, IX и XII в опухолях головного мозга. Раков 12:1723. doi: 10.3390/cancers12071723
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Хачински В., Эйнхаупл К., Гантен Д., Аллади С., Брейн К., Стефан Б.К.М. и др. (2019). Предотвращение деменции путем предотвращения инсульта: Берлинский манифест. Альцгеймер Демент. 15, 961–984. дои: 10.1016/j.jalz.2019.06.001
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Hall, C.N., Reynell, C., Gesslein, B. , Hamilton, N.B., Mishra, A., Sutherland, B.A., et al. (2014). Капиллярные перициты регулируют мозговой кровоток в норме и при патологии. Природа 508, 55–60. doi: 10.1038/nature13165
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Хан М., Квон И., Ха Дж., Ким Дж., Ча М.Дж., Ким Ю.Д. и др. (2020). Лечение коллатеральной аугментации комбинацией ацетазоламида и наклона головы вниз на модели ишемического инсульта у крыс. Дж. Клин. Неврологи. 73, 252–258. doi: 10.1016/j.jocn.2020.01.079
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Эрнандес-Гильямон, М., Мавирт, С., Фоссати, С., Блейс, С., Парес, М., Пенальба, А., и др. (2010). Матриксная металлопротеиназа 2 (ММР-2) расщепляет растворимые васкулотропные бета-амилоидные мутанты E22Q и L34V, задерживая их токсичность для эндотелиальных клеток микрососудов головного мозга человека. J. Biol. хим. 285, 27144–27158. doi: 10.1074/jbc.M110.135228
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Хилво М. , Баранаускене Л., Салцано А. М., Скалони А., Матулис Д., Инноченти А. и др. (2008). Биохимическая характеристика СА IX, одного из наиболее активных изоферментов карбоангидразы. J. Biol. хим. 283, 27799–27809. doi: 10.1074/jbc.M800938200
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Хун, Дж. Х., Мухаммад, Э., Чжэн, К., Хершковитц, Э., Алкринави, С., Ловенталь, Н., и др. (2015). Существенная роль карбоангидразы XII в секреторной жидкости железы и секреции HCO3 (-), выявленная мутацией, вызывающей заболевание человека. Дж. Физиол. 593, 5299–5312. doi: 10.1113/JP271378
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ховарт К., Сазерленд Б., Чой Х. Б., Мартин К., Линд Б. Л., Хеннуф Л. и др. (2017). Критическая роль астроцитов в гиперкапнической вазодилатации головного мозга. J. Neurosci. 37, 2403–2414. doi: 10.1523/JNEUROSCI.0005-16.2016
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ху, М. Ю., Линь, Ю.Ю., Чжан, Б.Дж., Лу, Д.Л., Лу, З.К., и Цай, В. (2019). Обновление активации воспаления в микроглии/макрофагах при старении и связанных со старением заболеваниях. ЦНС Неврологи. тер. 25, 1299–1307. doi: 10.1111/cns.13262
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Хуан Б.Р., Лю Ю.С., Лай С.В., Линь Х.Дж., Шен С.К., Ян Л.Ю. и др. (2020). CAIX регулирует подвижность ГБМ, адгезию и поляризацию ТАМ через EGFR/STAT3 в условиях гипоксии. Междунар. Дж. Мол. науч. 21:5838. дои: 10.3390/ijms21165838
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Хуанг П. Л., Хуанг З., Машимо Х., Блох К. Д., Московиц М. А., Беван Дж. А. и др. (1995). Гипертония у мышей, у которых отсутствует ген эндотелиальной синтазы оксида азота. Природа 377, 239–242. doi: 10.1038/377239a0
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Huneau, C., Benali, H., and Chabriat, H. (2015). Исследование нейроваскулярной связи человека с использованием функциональной нейровизуализации: критический обзор динамических моделей. Фронт. Неврологи. 9:467. doi: 10.3389/fnins.2015.00467
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ядекола, К. (2010). Перекрытие нейродегенеративных и сосудистых факторов в патогенезе деменции. Акта Нейропатол. 120, 287–296. doi: 10.1007/s00401-010-0718-6
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ядекола, К. (2017). Нейроваскулярная единица взросления: путешествие через нервно-сосудистую связь в здоровье и болезни. Нейрон 96, 17–42. doi: 10.1016/j.neuron.2017.07.030
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ядекола, К., и Недергаард, М. (2007). Глиальная регуляция мозгового микроциркуляторного русла. Нац. Неврологи. 10, 1369–1376. doi: 10.1038/nn2003
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Илиес М. , Банчу М. Д., Илиес М. А., Скоццафава А., Капрою М. Т. и Супуран С. Т. (2002). Активаторы карбоангидразы: разработка высокоаффинных изоферментов I, II и IV, включающих фрагменты три-/тетразамещенного пиридиний-азола. J. Med. хим. 45, 504–510. doi: 10.1021/jm011031n
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Илиес М. А., Масереэль Б., Ролин С., Скоццафава А., Кампеану Г., Кимпеану В. и др. (2004). Ингибиторы карбоангидразы: ароматические и гетероциклические сульфаниламиды, включающие адамантиловые фрагменты, обладающие сильным противосудорожным действием. Биоорг. Мед. хим. 12, 2717–2726. doi: 10.1016/j.bmc.2004.03.008
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Илиес, М. А., Вулло, Д., Пасторек, Дж., Скоццафава, А., Илиес, М., Капрою, М. Т., и соавт. (2003). Ингибиторы карбоангидразы. Ингибирование ассоциированного с опухолью изофермента IX производными галогенсульфаниламидов и галогенофениламинобензоламидов. J. Med. хим. 46, 2187–2196. doi: 10.1021/jm021123s
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Илиес, М. А., и Винум, Дж.-Ю. (2019). «Глава 16. Ингибиторы карбоангидразы для лечения опухолей: терапевтические, иммунологические и диагностические инструменты, нацеленные на изоформы IX и XII» в Карбоангидразы. eds CT Supuran and A. Nocentini (США: Academic Press). 331–365. doi: 10.1016/B978-0-12-816476-1.00016-2
Полный текст CrossRef | Google Scholar
Илифф Дж. Дж., Ван М., Ляо Ю., Плогг Б. А., Пэн В., Гундерсен Г. А. и др. (2012). Параваскулярный путь способствует оттоку спинномозговой жидкости через паренхиму головного мозга и клиренсу интерстициальных растворенных веществ, включая бета-амилоид. наук. Перевод Мед. 4, 147ра111. doi: 10.1126/scitranslmed.3003748
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Имтаияз Хассан М., Шаджи Б., Вахид А., Ахмад Ф. и Слай В. С. (2013). Структура, функция и применение изоферментов карбоангидразы. Биоорг. Мед. хим. 21, 1570–1582 гг. doi: 10.1016/j.bmc.2012.04.044
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Иванов С., Ляо С. Ю., Иванова А., Данилкович-Мягкова А., Тарасова Н., Вейрих Г. и др. (2001). Экспрессия индуцируемых гипоксией трансмембранных карбоангидраз клеточной поверхности при раке человека. 916:50 утра. Дж. Патол. 158, 905–919. doi: 10.1016/S0002-9440(10)64038-2
Полный текст CrossRef | Google Scholar
Иванов С.В., Кузьмин И., Вей М.-Х., Пак С., Гейл Л., Джонсон Б.Е. и др. (1998). Понижающая регуляция трансмембранных карбоангидраз в клеточных линиях почечно-клеточной карциномы трансгенами фон Хиппеля-Линдау дикого типа. Проц. Натл. акад. науч. 95:12596. doi: 10.1073/pnas.95.21.12596
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Цзян, К.Т., Ву, В.Ф., Дэн, Ю.Х., и Ге, Дж.В. (2020). Модуляторы активации и поляризации микроглии при ишемическом инсульте (обзор). мол. Мед. Респ. 21, 2006–2018. doi: 10.3892/mmr.2020.11003
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Цзяо Х., Ван З., Лю Ю., Ван П. и Сюэ Ю. (2011). Специфическая роль белков плотного контакта клаудина-5, окклюдина и ZO-1 гематоэнцефалического барьера при очаговом церебральном ишемическом инсульте. Дж. Мол. Неврологи. 44, 130–139. doi: 10.1007/s12031-011-9496-4
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Джонсон, Б.А. (2005). Недавние успехи в разработке методов лечения алкогольной и кокаиновой зависимости: основное внимание уделяется топирамату и другим модуляторам функции ГАМК или глутамата. Препараты ЦНС 19, 873–896. doi: 10.2165/00023210-200519100-00005
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Джоши А. У., Минхас П. С., Лидделоу С. А., Хайлеселассие Б., Андреассон К. И. и Дорн Г. В. II и др. (2019). Фрагментированные митохондрии, высвобождаемые из микроглии, запускают астроцитарный ответ А1 и распространяют воспалительную нейродегенерацию. Нац. Неврологи. 22, 1635–1648. doi: 10.1038/s41593-019-0486-0
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Кан М. и Яо Ю. (2020). Изменения базальной мембраны при ишемическом инсульте. Инсульт 51, 1344–1352. doi: 10.1161/STROKEAHA.120.028928
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Каплан, Л., Чоу, Б.В., и Гу, К. (2020). Нейрональная регуляция гематоэнцефалического барьера и нейроваскулярная связь. Нац. Преподобный Нейроски. 21, 416–432. doi: 10.1038/s41583-020-0322-2
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Кида Э., Пальминелло С., Голабек А. А., Валус М., Вежба-Бобровиц Т., Рабе А. и др. (2006). Карбоангидраза II в развивающемся и взрослом мозге человека. J. Невропатология. Эксп. Нейрол. 65, 664–674. дои: 10.1097/01.jnen.0000225905.52002.3e
CrossRef Полный текст | Google Scholar
Ким С. Х., Ан Дж. Х., Ян Х. , Ли П., Кох Г. Ю. и Чон Ю. (2020). Церебральная амилоидная ангиопатия усугубляет нарушение периваскулярного клиренса в мышиной модели болезни Альцгеймера. Акта Нейропатол. коммун. 8:181. doi: 10.1186/s40478-020-01042-0
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Кисс Б., Даллинджер С., Финдл О., Райнер Г., Эйхлер Х. Г. и Шметтерер Л. (1999). Вызванная ацетазоламидом церебральная и глазная вазодилатация у людей не зависит от оксида азота. утра. Дж. Физиол. 276, R1661–R1667. doi: 10.1152/ajpregu.1999.276.6.R1661
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Кнопман Д.С., Джонс Д.Т. и Грейциус М.Д. (2021). Неспособность продемонстрировать эффективность адуканумаба: анализ исследований EMERGE и ENGAGE по данным Biogen. Декабрь 2019. Альцгеймер Демент. 17, 696–701. doi: 10.1002/alz.12213
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Кришнамурти В. М., Кауфман Г. К., Урбах А. Р., Гитлин И., Гудиксен К. Л., Вейбель Д. Б. и др. (2008). Карбоангидраза как модель для биофизических и физико-органических исследований белков и связывания белок-лиганд. Хим. Ред. 108, 946–1051. doi: 10.1021/cr050262p
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Куриакосе Д. и Сяо З. (2020). Патофизиология и лечение инсульта: современное состояние и перспективы на будущее. Междунар. Дж. Мол. науч. 21:7609. doi: 10.3390/ijms21207609
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Кузьма Э., Лурида И., Мур С. Ф., Левин Д. А., Укумунне О. К. и Ллевеллин Д. Дж. (2018). Риск инсульта и деменции: систематический обзор и метаанализ. Альцгеймер Демент. 14, 1416–1426. doi: 10.1016/j.jalz.2018.06.3061
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Li, M., Wang, W., Mai, H., Zhang, X., Wang, J., Gao, Y., et al. (2016). Метазоламид улучшает неврологическое поведение путем ингибирования апоптоза нейронов у мышей с субарахноидальным кровоизлиянием. г. научн. Респ. 6:35055. doi: 10.1038/srep35055
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Liddelow, S.A., Guttenplan, K.A., Clarke, L.E., Bennett, F.C., Bohlen, C.J., Schirmer, L., et al. (2017). Нейротоксические реактивные астроциты индуцируются активированной микроглией. Природа 541, 481–487. doi: 10.1038/nature21029
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Лю К., Вэй Ю., Ван Дж., Пи Л., Хуанг Дж. и Ван П. (2012). Аутоантитела к карбоангидразам III и IV при ревматоидном артрите, системной красной волчанке, сахарном диабете, гипертонической болезни почек и сердечной недостаточности. клин. Дев. Иммунол. 2012:354594. doi: 10.1155/2012/354594
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Лоххед Дж. Дж., Маккаффри Г., Куигли К. Э., Финч Дж., Демарко К. М., Наметц Н. и др. (2010). Окислительный стресс увеличивает проницаемость гематоэнцефалического барьера и вызывает изменения окклюдина во время гипоксии-реоксигенации. Дж. Цереб. Кровоток Метаб. 30, 1625–1636. doi: 10.1038/jcbfm.2010.29
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Лок Дж., Ван С. С., Син С. Х., Маки Т. К., Ву Л. М., Го С. З. и др. (2015). Ориентация на сосудисто-нервную единицу при травме головного мозга. ЦНС Неврологи. тер. 21, 304–308. doi: 10.1111/cns.12359
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Лоренцл С., Альберс Д. С., Релкин Н., Нгьюен Т., Хильгенберг С. Л., Чиричиньо Дж. и др. (2003). Повышение уровня матриксной металлопротеиназы-9 в плазме у пациентов с болезнью Альцгеймера. Нейрохим. Междунар. 43, 191–196. doi: 10.1016/S0197-0186(03)00004-4
Полный текст CrossRef | Google Scholar
Лу Х., Чжан Х. и Цзян Ю. (2020). Метазоламид при высотных болезнях. евро. Дж. Фарм. науч. 148:105326. doi: 10.1016/j.ejps.2020.105326
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Мэнли Г. Т., Фуджимура М., Ма Т., Ношита Н., Филиз Ф., Боллен А. В. и др. (2000). Делеция аквапорина-4 у мышей уменьшает отек головного мозга после острой водной интоксикации и ишемического инсульта. Нац. Мед. 6, 159–163. doi: 10.1038/72256
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Мао, С.Ю., Цао, Ю.Г., Цзи, З., Чжоу, Х.Х., Лю, З.К., и Сунь, Х.Л. (2015). Топирамат защищает от эксайтотоксичности глутамата посредством активации BDNF/TrkB-зависимого пути ERK в нейронах гиппокампа грызунов. Прог. Нейропсихофармакол. биол. Психиатрия 60, 11–17. doi: 10.1016/j.pnpbp.2015.01.015
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Марко С. и Скапер С. Д. (2006). Амилоидный бета-пептид 1-42 изменяет распределение и экспрессию белков плотных соединений в эндотелиальных клетках микрососудов головного мозга. Неврологи. лат. 401, 219–224. doi: 10.1016/j.neulet.2006.03.047
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Maren, TH (1967). Карбоангидраза: химия, физиология и ингибирование. Физиол. Ред. 47, 595–781. doi: 10.1152/physrev.1967.47.4.595
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Мбоге, М.Ю., Махон, Б.П., Маккенна, Р., и Фрост, С.К. (2018). Карбоангидразы: роль в контроле pH и раке. Метаболиты 8:19. doi: 10.3390/metabo8010019
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
McAlpine, C.S., Park, J., Griciuc, A., Kim, E., Choi, S.H., Iwamoto, Y., et al. (2021). Астроцитарный интерлейкин-3 программирует микроглию и ограничивает болезнь Альцгеймера. Природа 595, 701–706. doi: 10.1038/s41586-021-03734-6
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
МакКоннелл, Х.Л., Ли, З., Вольтьер, Р.Л., и Мишра, А. (2019). Дисфункция астроцитов и сосудисто-нервные нарушения при неврологических расстройствах: корреляция или причинно-следственная связь? Нейрохим. Междунар. 128, 70–84. doi: 10.1016/j.neuint.2019.04.005
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
McDonald, P.C., Chia, S., Bedard, P.L., Chu, Q., Lyle, M., Tang, L., et al. (2020). Исследование фазы 1 SLC-0111, нового ингибитора карбоангидразы IX, у пациентов с запущенными солидными опухолями. 916:50 утра. Дж. Клин. Онкол. 43, 484–490. doi: 10.1097/COC.0000000000000691
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Маккенна Р. и Супуран К. Т. (2014). Разработка лекарственных препаратов ингибиторов карбоангидразы. Субъячейка. Биохим. 75, 291–323. doi: 10.1007/978-94-007-7359-2_15
CrossRef Полный текст | Google Scholar
Местре Х., Мори Ю. и Недергаард М. (2020). Глимфатическая система мозга: текущие споры. Trends Neurosci. 43, 458–466. doi: 10.1016/j.tins.2020.04.003
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Mincione, F., Scozzafava, A., and Supuran, CT (2007). Разработка ингибиторов карбоангидразы местного действия в качестве средств против глаукомы. Курс. Верхний. Мед. хим. 7, 849–854. doi: 10.2174/156802607780636735
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Мишра, С. Б., Тивари, М., и Супуран, К. Т. (2020). Прогресс в разработке ингибиторов карбоангидразы человека и их фармакологических приложений: где мы находимся сегодня? Мед. Рез. Ред. 40, 2485–2565. doi: 10.1002/med.21713
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Мохамед И. Н., Ишрат Т., Фаган С. К. и Эль-Ремесси А. Б. (2015). Роль активации инфламмасом в патофизиологии сосудистых заболеваний сосудисто-нервного блока. Антиоксидант. Окислительно-восстановительный сигнал. 22, 1188–1206. doi: 10.1089/ars.2014.6126
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Монтанер Дж., Рамиро Л., Симатс А., Эрнандес-Гильямон М., Дельгадо П., Бустаманте А. и др. (2019). Матриксные металлопротеиназы и ADAM при инсульте. Сотовый. Мол. Жизнь наук. 76, 3117–3140. doi: 10.1007/s00018-019-03175-5
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Мори К., Огава Ю., Эбихара К., Тамура Н., Таширо К., Кувахара Т. и др. (1999). Выделение и характеристика CA XIV, новой мембраносвязанной карбоангидразы из почек мышей. J. Biol. хим. 274, 15701–15705. doi: 10.1074/jbc.274.22.15701
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Morikawa, E., Moskowitz, M.A., Huang, Z., Yoshida, T., Irikura, K., and Dalkara, T. (1994). Инфузия L-аргинина способствует зависимой от оксида азота вазодилатации, увеличивает регионарный мозговой кровоток и уменьшает объем инфаркта у крыс. Инсульт 25, 429–435. doi: 10.1161/01.STR.25.2.429
CrossRef Полный текст | Google Scholar
Моррис, А. В., Караре, Р. О., Шрайбер, С., и Хоукс, К. А. (2014). Цереброваскулярная базальная мембрана: роль в клиренсе β-амилоида и церебральной амилоидной ангиопатии. Фронт. Стареющие нейроски. 6:251. doi: 10.3389/fnagi.2014.00251
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Недергаард, М., и Голдман, С.А. (2020). Глимфатическая недостаточность как конечный общий путь к деменции. Наука 370, 50–56. doi: 10.1126/science.abb8739
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Нисимори И., Минакучи Т., Ониши С., Вулло Д., Скоццафава А. и Супуран С. Т. (2007). Ингибиторы карбоангидразы. Клонирование ДНК, характеристика и исследования ингибирования секреторной изоформы VI человека, новой мишени для сульфонамидных и сульфаматных ингибиторов. J. Med. хим. 50, 381–388. doi: 10.1021/jm0612057
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Нисимори И., Вулло Д., Инноченти А., Скоццафава А., Мастролоренцо А. и Супуран К. Т. (2005). Ингибиторы карбоангидразы. Митохондриальный изофермент VB как новая мишень для сульфонамидных и сульфаматных ингибиторов. J. Med. хим. 48, 7860–7866. doi: 10.1021/jm050483n
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Нортли, Р., Корте, Н., Искьердо, П., Хирунпаттарасилп, К., Мишра, А., Яунмуктане, З., и соавт. (2019). Олигомеры β-амилоида сужают капилляры человека при болезни Альцгеймера посредством передачи сигналов перицитам. Наука 365:eaav9518. doi: 10.1126/science.aav9518
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Окль П., Хальбгебауэр С., Андерл-Штрауб С., Штайнакер П., Хусс А. М. и Нойгебауэр Х. (2019). Глиальный фибриллярный кислый белок в сыворотке увеличивается при болезни Альцгеймера и коррелирует с когнитивными нарушениями. Дж. Альцгеймер Дис. 67, 481–488. doi: 10.3233/JAD-180325
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Оджо Дж. О., Рид Дж. М., Крайнен Г., Валлабханени П., Эванс Дж., Шеклтон Б. и др. (2021). Молекулярная патобиология сосудов головного мозга при старении и в случаях болезни Альцгеймера с церебральной амилоидной ангиопатией. Фронт. Стареющие нейроски. 13:658605. doi: 10.3389/fnagi.2021.658605
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Окудаира Ю., Бандох К., Араи Х. и Сато К. (1995). Оценка ацетазоламидного теста. Вазореактивность и объем мозговой крови. Инсульт 26, 1234–1239. doi: 10.1161/01.STR.26.7.1234
CrossRef Полный текст | Google Scholar
Олдендорф, У. Х., Корнфорд, М. Е., и Браун, У. Дж. (1977). Большая кажущаяся работоспособность гематоэнцефалического барьера: исследование митохондриального состава капиллярных эндотелиальных клеток головного мозга и других тканей крысы. г. н.э. Нейрол. 1, 409–417. doi: 10.1002/ana.410010502
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Пан П., Леппилампи М., Пасторекова С., Пасторек Дж., Вахид А., Слай В. С. и др. (2006). Экспрессия гена карбоангидразы у мышей с дефицитом CA II (Car2-/-) и дефицитом CA IX (Car9-/-). J. Physiol. 571, 319–327. doi: 10.1113/jphysiol.2005.102590
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Пан П.В., Парккила А.К., Аутио С., Хилво М., Сормунен Р., Пасторекова С. и др. (2012). Фенотип мозга мышей с дефицитом карбоангидразы IX. трансгенных рез. 21, 163–176. doi: 10.1007/s11248-011-9520-z
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Пароди-Руллан Р., Гисо Дж., Кабрера Э., Ростаньо А. и Фоссати С. (2020). Гетерогенные виды бета-амилоида болезни Альцгеймера по-разному влияют на жизнеспособность эндотелиальных клеток головного мозга, целостность гематоэнцефалического барьера и ангиогенез. Стареющая ячейка. 19:e13258. doi: 10.1111/acel.13258
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Пароди-Руллан Р., Соне Дж. Ю. и Фоссати С. (2019). Эндотелиальная митохондриальная дисфункция при церебральной амилоидной ангиопатии и болезни Альцгеймера. J. Alzheimers Dis. 72, 1019–1039. doi: 10.3233/JAD-1
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Пасторек Дж. и Пасторекова С. (2015). Карбоангидраза IX, индуцированная гипоксией, как мишень для терапии рака: от биологии к клиническому применению. Семин. Рак биол. 31, 52–64. doi: 10.1016/j.semcancer.2014.08.002
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Патрик П., Прайс Т. О., Диого А. Л., Шейбани Н., Бэнкс В. А. и Шах Г. Н. (2015). Топирамат защищает перициты от глюкозотоксичности: роль митохондриальной CA VA в цереброваскулярных заболеваниях при диабете. Дж. Эндокринол. Диабет 2, 1–7. doi: 10.15226/2374-6890/2/2/00123
CrossRef Full Text | Google Scholar
Патрикайнен М., Пан П., Кулесская Н., Войкар В. и Парккила С. (2014). Роль карбоангидразы VI в восприятии горького вкуса: данные Car6 – / – модель мыши . Дж. Биомед. науч. 21, 82. doi: 10.1186/s12929-014-0082-2
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Павич Л. , Грюнвальд Ф., Райхманн К., Хорн Р., Китшенберг А., Хартманн А. и др. (1999). Однофотонная эмиссионная томография регионарного мозгового кровотока с 99mTc-HMPAO и тест с ацетазоламидом в оценке сосудистой и альцгеймеровской деменции. евро. Дж. Нукл. Мед. 26, 239–245. дои: 10.1007/s0025
383
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Пекни М. и Лейн Э. Б. (2007). Промежуточные филаменты и стресс. Опыт. Сотовый рез. 313, 2244–2254. doi: 10.1016/j.yexcr.2007.04.023
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Поллард А., Шепард Ф., Фрид Дж., Лидделл С. и Чакрабарти Л. (2016). Митохондриальное протеомное профилирование выявляет увеличение карбоангидразы II при старении и нейродегенерации. Старение 8, 2425–2436. doi: 10.18632/aging.101064
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Прайс, Т. О., Еранки, В., Бэнкс, В. А., Эркал, Н., и Шах, Г. Н. (2012). Лечение топираматом защищает перициты гематоэнцефалического барьера от вызванного гипергликемией окислительного повреждения у мышей с диабетом. Эндокринология 153, 362–372. doi: 10.1210/en.2011-1638
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Прайс, Т. О., Шейбани, Н., и Шах, Г. Н. (2017). Регуляция высокого индуцируемого глюкозой апоптоза перицитов головного мозга митохондриальной CA VA: специфическая мишень для профилактики диабетической цереброваскулярной патологии. Биохим. Биофиз. Акта Мол. Основа Дис. 1863, 929–935. doi: 10.1016/j.bbadis.2017.01.025
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Proescholdt, M.A., Mayer, C., Kubitza, M., Schubert, T., Liao, S.-Y., Stanbridge, E.J., et al. (2005). Экспрессия индуцируемых гипоксией карбоангидраз в опухолях головного мозга. Нейроонкология 7, 465–475. doi: 10.1215/S1152851705000025
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Provensi, G. , Carta, F., Nocentini, A., Supuran, C.T., Casamenti, F., Passani, M.B., et al. (2019). Новичок в блоке? Карбоангидразы как возможные новые мишени при болезни Альцгеймера. Int J Mol Sci 20:4724. doi: 10.3390/ijms20194724
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Пун, П. Б., Лу, Дж., и Мучхала, С. (2009). Участие АФК в дисфункции ГЭБ. Свободный радикал. Рез. 43, 348–364. doi: 10.1080/10715760
1902
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Пуркерсон, Дж. М., и Шварц, Г. Дж. (2007). Роль карбоангидразы в физиологии почек. Почки Междунар. 71, 103–115. doi: 10.1038/sj.ki.5002020
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Quintana, D.D., Anantula, Y., Garcia, J.A., Engler-Chiurazzi, E.B., Sarkar, S.N., Corbin, D.R., et al. (2021). Дегенерация микрососудов происходит до появления бляшек и прогрессирует с возрастом у мышей 3xTg AD. Нейробиол. Старение 105, 115–128. doi: 10.1016/j.neurobiolaging.2021.04.019
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Раденович Л., Ненадич М., Уламек-Козиол М., Янушевский С., Чучвар С. Дж., Андьюс П. Р. и др. (2020). Неоднородность в распределении активированной микроглии и астроцитов в головном мозге в ишемической модели болезни Альцгеймера у крыс после 2 лет выживания. Старение 12, 12251–12267. doi: 10.18632/aging.103411
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Расмуссен, Дж. К., и Боэдткьер, Э. (2018). Ингибиторы карбоангидразы модифицируют переходные процессы внутриклеточного pH и сокращения средних мозговых артерий крыс во время CO. Дж. Цереб. Кровоток Метаб. 38, 492–505. doi: 10.1177/0271678X17699224
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Риз Т.С. и Карновский М.Дж. (1967). Тонкая структурная локализация гематоэнцефалического барьера для экзогенной пероксидазы. Дж. Сотовый. биол. 34, 207–217. doi: 10.1083/jcb.34.1.207
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ричи, Н.Д., Багготт, А.В., и Эндрю Тодд, В.Т. (2012). Ацетазоламид для профилактики острой горной болезни – систематический обзор и метаанализ. J. Travel Med. 19, 298–307. doi: 10.1111/j.1708-8305.2012.00629.x
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Рассел С. М., Ву Х. Х., Силлер К., Панаски Д. и Леру П. Д. (2008). Оценка резерва коллатерального кровотока средней мозговой артерии с помощью транскраниальной допплерографии с ацетазоламидом у пациентов с окклюзионной болезнью сонных артерий. Хирург. Нейрол. 70, 466–470. doi: 10.1016/j.surneu.2007.10.030
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Ruusuvuori, E., Huebner, A.K., Kirilkin, I., Yukin, A.Y., Blaesse, P., Helmy, M., et al. (2013). Нейрональная карбоангидраза VII обеспечивает ГАМКергическую возбуждающую активность, усиливающую фебрильные судороги. Эмбо Дж. 32, 2275–2286. doi: 10.1038/emboj.2013.160
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Руусувуори Э. и Кайла К. (2014). Карбоангидразы и рН головного мозга в контроле возбудимости нейронов. Субъячейка. Биохим. 75, 271–290. doi: 10.1007/978-94-007-7359-2_14
CrossRef Full Text | Google Scholar
Руусувуори Э., Ли Х., Хутту К., Палва Дж. М., Смирнов С., Ривера К. и др. (2004). Изоформа карбоангидразы VII действует как молекулярный переключатель в развитии синхронного гамма-излучения пирамидных клеток CA1 гиппокампа. J. Neurosci. 24, 2699–2707. doi: 10.1523/JNEUROSCI.5176-03.2004
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Saarnio, J., Parkkila, S., Parkkila, A.K., Waheed, A., Casey, M.C., Zhou, X.Y., et al. (1998). Иммуногистохимия изофермента карбоангидразы IX (MN/CA IX) в кишечнике человека выявляет поляризованную экспрессию в эпителиальных клетках с самой высокой пролиферативной способностью. J. Histochem. Цитохим. 46, 497–504. doi: 10.1177/002215549804600409
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Сакко Р. Л., Каснер С. Е., Бродерик Дж. П., Каплан Л. Р., Коннорс Дж. Дж., Кулебрас А. и др. (2013). Обновленное определение инсульта для 21 века: заявление для медицинских работников Американской кардиологической ассоциации/Американской ассоциации инсульта. Инсульт 44, 2064–2089. doi: 10.1161/STR.0b013e318296aeca
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Сагаре А.П., Белл Р.Д., Чжао З., Ма К., Винклер Э.А., Раманатан А. и др. (2013). Потеря перицитов влияет на альцгеймероподобную нейродегенерацию у мышей. Нац. коммун. 4:2932. doi: 10.1038/ncomms3932
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Саламе Т. С., Мортелл В. Г., Логсдон А. Ф., Баттерфилд Д. А. и Бэнкс В. А. (2019 г.)). Нарушение гиппокампального и гипоталамического гематоэнцефалического барьера в модели диабета II типа, вызванной диетой, с ожирением: профилактика и лечение ингибитором митохондриальной карбоангидразы топираматом. Жидкостные барьеры CNS 16:1. doi: 10.1186/s12987-018-0121-6
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Саламе Т. С., Шах Г. Н., Прайс Т. О., Хайден М. Р. и Бэнкс В. А. (2016). Нарушение гематоэнцефалического барьера и дисфункция нейроваскулярных единиц у мышей с диабетом: защита с помощью ингибитора митохондриальной карбоангидразы топирамата. Дж. Фармакол. Эксп. тер. 359, 452–459. doi: 10.1124/jpet.116.237057
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Санку Р.К.К., Джон Дж.С., Сальковиц М., Илиес М.А. и Уокер Э.А. (2018). Потенциальные разрушители и усилители обучения и памяти в простой однодневной оперантной задаче у мышей. Поведение. Фармакол. 29, 482–492. doi: 10.1097/FBP.0000000000000400
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Сарвари С., Моакеди Ф., Хон Э., Симпкинс Дж. В. и Рен X. (2020). Механизмы открытия гематоэнцефалического барьера и метаболической цереброваскулярной ишемии с акцентом на ишемический инсульт. Метаб. Мозг Дис. 35, 851–868. doi: 10.1007/s11011-020-00573-8
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Савва Г. М. и Стефан Б. К. (2010). Эпидемиологические исследования влияния инсульта на деменцию: систематический обзор. Ход 41, e41–46. doi: 10.1161/STROKEAHA.109.559880
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Шмидт С. Д., Коста А., Рани Б., Годфрид Нахтигал Э., Пассани М. Б., Карта Ф. и др. (2020). Роль карбоангидразы в угасании контекстуальной памяти о страхе. г. Проц. Натл. акад. науч. США 117, 16000–16008. doi: 10.1073/pnas.19106
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Schreibelt, G., Kooij, G., Reijerkerk, A., Van Doorn, R., Gringhuis, S.I., Van Der Pol, S., et al. (2007). Активные формы кислорода изменяют динамику плотных соединений эндотелия головного мозга через RhoA. киназа PI3 и передача сигналов PKB. FASEB J. 21, 3666–3676. doi: 10.1096/fj.07-8329com
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Scozzafava, A., Briganti, F., Ilies, M.A., and Supuran, C.T. (2000). Ингибиторы карбоангидразы: синтез мембранонепроницаемых низкомолекулярных сульфаниламидов, обладающих селективностью in vivo в отношении мембраносвязанных изоферментов по сравнению с цитозольными. J. Med. хим. 43, 292–300. doi: 10.1021/jm9+
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Scozzafava, A., Supuran, C.T., and Carta, F. (2013). Ингибиторы карбоангидразы против ожирения: обзор литературы и патентов. Эксперт. мнение тер. Пат. 23, 725–735. doi: 10.1517/13543776.2013.7
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Сечкин Х., Симшек С., Озтюрк Э., Йигитканли К., Озен О., Бешалти О. и др. (2009). Топирамат ослабляет повреждение гиппокампа после экспериментального субарахноидального кровоизлияния у кроликов. Нейрол. Рез. 31, 490–495. doi: 10.1179/016164108X339369
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Секердаг Э., Солараглу И. и Гурсой-Оздемир Ю. (2018). Механизмы гибели клеток при инсульте и новые варианты молекулярного и клеточного лечения. Курср. Нейрофармакол. 16, 1396–1415. doi: 10.2174/1570159X16666180302115544
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Севиньи Дж., Чиао П., Бюссьер Т., Вайнреб П. Х., Уильямс Л., Майер М. и др. (2017). Приложение: антитело адуканумаб уменьшает бляшки Абета при болезни Альцгеймера. Природа 546:564. doi: 10.1038/nature22809
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Шабана, А. М., и Илиес, Массачусетс (2019). Доставка лекарств к гипоксическим опухолям, нацеленным на карбоангидразу IX. США: Американское химическое общество. 223–252. doi: 10.1021/bk-2019-1309.ch010
Полный текст CrossRef | Google Scholar
Shah, G. N., Hewett-Emmett, D., Grubb, J. H., Migas, M. C., Fleming, R. E., Waheed, A., et al. (2000). Митохондриальная карбоангидраза CA VB: различия в тканевом распределении и характере эволюции по сравнению с CA VA предполагают разные физиологические роли. Проц. Натл. акад. науч. США 97, 1677–1682. doi: 10.1073/pnas.97.4.1677
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Шах Г. Н., Морофуджи Ю., Бэнкс В. А. и Прайс Т. О. (2013a). Индуцированное высоким содержанием глюкозы митохондриальное дыхание и активные формы кислорода в перицитах головного мозга мышей обращаются фармакологическим ингибированием митохондриальных карбоангидраз: последствия для церебральных микрососудистых заболеваний при диабете. Биохим. Биофиз. Рез. коммун. 440, 354–358. doi: 10.1016/j.bbrc.2013.09.086
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Shah, G. N., Rubbelke, T. S., Hendin, J., Nguyen, H., Waheed, A., Shoemaker, J. D., et al. (2013б). Направленный мутагенез митохондриальных карбоангидраз VA и VB вовлекает оба фермента в детоксикацию аммиака и метаболизм глюкозы. Проц. Натл. акад. науч. США 110, 7423–7428. doi: 10.1073/pnas.1305805110
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Шах Г. Н., Улмасов Б., Вахид А., Беккер Т., Макани С., Свичар Н. и др. (2005). Мыши с нокаутом карбоангидразы IV и XIV: роль соответствующих карбоангидраз в буферизации внеклеточного пространства головного мозга. г. Проц. Натл. акад. науч. США 102, 16771–16776. doi: 10.1073/pnas.0508449102
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Шин, Дж. А., Юн, Дж. К., Ким, М., и Пак, Э. М. (2016). Активация классических подтипов рецепторов эстрогена уменьшает нарушение плотных контактов эндотелиальных клеток головного мозга при ишемическом/реперфузионном повреждении. Свободный радикал. биол. Мед. 92, 78–89. doi: 10.1016/j.freeradbiomed. 2016.01.010
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Сильверман, Д. Н., и Линдског, С. (1988). Каталитический механизм карбоангидразы: последствия ограничивающего скорость протолиза воды. Согл. хим. Рез. 21, 30–36. doi: 10.1021/ar00145a005
CrossRef Full Text | Google Scholar
Слай В.С. и Ху П.Ю. (1995). Карбоангидразы человека и недостаточность карбоангидразы. год. Преподобный Биохим. 64, 375–401. doi: 10.1146/annurev.bi.64.070195.002111
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Smaine, F.Z., Winum, J.Y., Montero, J.L., Regainia, Z., Vullo, D., Scozzafava, A., et al. (2007). Ингибиторы карбоангидразы: селективное ингибирование внеклеточных опухолевых изоформ IX и XII по сравнению с изоферментами I и II гликозилтиоуреидосульфонамидами. Биоорг. Мед. хим. лат. 17, 5096–5100. doi: 10.1016/j.bmcl.2007.07.019
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Смит А. Дж., Дуан Т. и Веркман А. С. (2019). Аквапорин-4 уменьшает невропатологию в мышиной модели болезни Альцгеймера путем ремоделирования структуры астроцитов вокруг бляшек. Acta Neuropathol. коммун. 7:74. doi: 10.1186/s40478-019-0728-0
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Солесио, М. Э., Пейшото, П. М., Дебуре, Л., Мадамба, С. М., Де Леон, М. Дж., Вишневски, Т., и др. (2018). Ингибирование карбоангидразы избирательно предотвращает нейроваскулярную митохондриальную токсичность амилоида β. Стареющая ячейка. 17:e12787. doi: 10.1111/acel.12787
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Сотолонго, К., Гизо, Дж., и Ростаньо, А. (2020). Активация Nrf2 через ось PI3K/GSK-3 защищает нейронные клетки от Abeta-опосредованного окислительного и метаболического повреждения. Альцгеймер Res. тер. 12:13. doi: 10.1186/s13195-019-0578-9
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Стэмс Т. , Наир С.К., Окуяма Т., Вахид А., Слай У.С. и Кристиансон Д.В. (1996). Кристаллическая структура секреторной формы связанной с мембраной карбоангидразы IV человека с разрешением 2,8 Å. Проц. Натл. акад. науч. США 93, 13589–13594. doi: 10.1073/pnas.93.24.13589
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Stoppe, G., Schutze, R., Kogler, A., Staedt, J., Munz, D.L., Emrich, D., et al. (1995). Цереброваскулярная реактивность на ацетазоламид при (сенильной) деменции альцгеймеровского типа: связь с тяжестью заболевания. Деменция 6, 73–82. doi: 10.1159/000106925
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Supuran, CT (2008). Карбоангидразы: новые терапевтические применения ингибиторов и активаторов. Нац. Преподобный Друг Дисков. 7, 168–181. дои: 10.1038/nrd2467
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Supuran, CT (2011). Ингибиторы и активаторы карбоангидразы для новых терапевтических применений. Будущее Мед. хим. 3, 1165–1180. doi: 10.4155/fmc.11.69
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Supuran, CT (2012). Ингибиторы карбоангидразы как перспективные препараты для лечения ожирения. Экспертное заключение. Эмердж. Наркотики 17, 11–15. дои: 10.1517/14728214.2012.664132
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Supuran, CT (2018). Применение ингибиторов карбоангидразы при заболеваниях почек и центральной нервной системы. Экспертное заключение. тер. Пат. 28, 713–721. doi: 10.1080/13543776.2018.1519023
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Супуран, К.Т., и Капассо, К. (2020). Антибактериальные ингибиторы карбоангидразы: обновленная информация о последних публикациях. Экспертное заключение. тер. Пат. 30, 963–982. doi: 10.1080/13543776.2020.1811853
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Супуран, К. Т., Скоццафава, А., и Казини, А. (2003). Ингибиторы карбоангидразы. Мед. Рез. 23, 146–189. doi: 10.1002/med.10025
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Supuran, C.T., Scozzafava, A., Ilies, M.A., Iorga, B., Cristea, T., Briganti, F., et al. (1998). Ингибиторы карбоангидразы — Часть 53. Синтез замещенных пиридиниевых производных ароматических сульфаниламидов: первые неполимерные мембранонепроницаемые ингибиторы с селективностью по изоферменту IV. евро. Дж. Мед. хим. 33, 577–594. doi: 10.1016/S0223-5234(98)80017-2
Полный текст CrossRef | Google Scholar
Супуран, К.Т., и Винум, Дж.Ю. (2015). Ингибиторы карбоангидразы IX в терапии рака: обновление. Будущее Мед. хим. 7, 1407–1414. doi: 10.4155/fmc.15.71
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Sweeney, MD, Montagne, A., Sagare, A.P., Nation, D.A., Schneider, L.S., Chui, H.C., et al. (2019а). Сосудистая дисфункция — игнорируемый партнер болезни Альцгеймера. Демент Альцгеймера. 15, 158–167.
Google Scholar
Суини, М. Д., Чжао, З., Монтань, А., Нельсон, А. Р., и Злокович, Б. В. (2019b). Гематоэнцефалический барьер: от физиологии к болезни и обратно. Физиол. Ред. 99, 21–78. doi: 10.1152/physrev.00050.2017
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Тан М., Гао Г., Руэда С. Б., Ю Х., Тибодо Д. Н., Авано Т. и др. (2017). Дефекты микроциркуляторного русла головного мозга и синдром дефицита Glut1 предотвращаются ранним восполнением белка переносчика глюкозы-1. Нац. коммун. 8:14152. doi: 10.1038/ncomms14152
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Танг Ю. и Ле В. (2016). Дифференциальные роли микроглии М1 и М2 при нейродегенеративных заболеваниях. мол. Нейробиол. 53, 1181–1194. doi: 10.1007/s12035-014-9070-5
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Тюттенберг Дж., Хейманн А. и Кемпски О. (2001). Оксид азота модулирует стимуляцию мозгового кровотока ацетазоламидом в коре головного мозга крыс: исследование лазерного допплеровского сканирования. Неврологи. лат. 315, 65–68. doi: 10.1016/S0304-3940(01)02325-4
Полный текст CrossRef | Google Scholar
Türeci, Ö, Sahin, U., Vollmar, E., Siemer, S., Göttert, E., Seitz, G., et al. (1998). Карбоангидраза человека XII: клонирование кДНК, экспрессия и хромосомная локализация гена карбоангидразы, который сверхэкспрессируется при некоторых видах почечно-клеточного рака. Проц. Натл. акад. науч. 95:7608. doi: 10.1073/pnas.95.13.7608
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Тернер, Р. Дж., и Шарп, Ф. Р. (2016). Влияние MMP9 на нарушение гематоэнцефалического барьера и геморрагическую трансформацию после ишемического инсульта. Фронт. Клетка. Неврологи. 10:56. doi: 10.3389/fncel.2016.00056
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Улланд, Т. К., и Колонна, М. (2018). TREM2 — ключевой игрок в биологии микроглии и болезни Альцгеймера. Нац. Преподобный Нейрол. 14, 667–675. doi: 10.1038/s41582-018-0072-1
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
ван Путтен, М., Фальке, К., Кафиц, К.В., Хофмейер, Дж., и Роуз, Ч.Р. (2021). Нарушение регуляции гомеостаза ионов астроцитов и его значение для повреждения головного мозга, вызванного инсультом. Междунар. Дж. Мол. науч. 22:5679. doi: 10.3390/ijms22115679
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Варгас Л. А., Пинилья О. А., Диас Р. Г., Сепульведа Д. Э., Свенсон Э. Р., Перес Н. Г. и др. (2016). Ингибиторы карбоангидразы снижают сердечную дисфункцию после длительной перевязки коронарных артерий у крыс. г. Сердечно-сосудистые заболевания. Патол. 25, 468–477. doi: 10.1016/j.carpath.2016.08.003
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Венкатешаппа, К. , Хариш, Г., Митри, Р. Б., Махадеван, А., Бхарат, М. М., и Шанкар, С. К. (2012). Повышенное окислительное повреждение и снижение антиоксидантной функции у стареющего человека в черной субстанции по сравнению с полосатым телом: последствия для болезни Паркинсона. Нейрохим. Рез. 37, 358–369. doi: 10.1007/s11064-011-0619-7
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Веркерке, М., Хол, Э.М., и Мидделдорп, Дж. (2021). Физиологическое и патологическое старение астроцитов головного мозга человека. Нейрохим. Рез. 46, 2662–2675. doi: 10.1007/s11064-021-03256-7
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Вильегас-Ллерена, К., Филлипс, А., Гарсия-Райтбок, П., Харди, Дж., и Покок, Дж. М. (2016). Гены микроглии, регулирующие нейровоспаление при прогрессировании болезни Альцгеймера. Курс. мнение Нейробиол. 36, 74–81. doi: 10.1016/j.conb.2015.10.004
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Вахид А. и Слай В. С. (2017). Карбоангидраза XII функционирует в норме и при болезни. Гена 623, 33–40. doi: 10.1016/j.gene.2017.04.027
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Wang X., Figueroa B. E., Stavrovskaya I. G., Zhang Y., Sirianni A. C., Zhu S., et al. (2009). Метазоламид и мелатонин ингибируют высвобождение митохондриального цитохрома С и оказывают нейропротекторное действие в экспериментальных моделях ишемического повреждения. г. Инсульт 40, 1877–1885 гг. doi: 10.1161/STROKEAHA.108.540765
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ван Х., Чжу С., Пей З., Дрозда М., Ставровская И. Г., Дель Синьор С. Дж. и др. (2008). Ингибиторы высвобождения цитохрома с с терапевтическим потенциалом при болезни Гентингтона. J. Neurosci. 28, 9473–9485. doi: 10.1523/JNEUROSCI.1867-08.2008
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Уиттингтон Д. А., Вахид А. , Ульмасов Б., Шах Г. Н., Грабб Дж. Х., Слай В. С. и др. (2001). Кристаллическая структура димерного внеклеточного домена карбоангидразы XII человека, битопного мембранного белка, сверхэкспрессируемого в некоторых раковых опухолевых клетках. г. Проц. Натл. акад. науч. США 98, 9545–9550. doi: 10.1073/pnas.161301298
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Уиллис, Э. Ф., Макдональд, К. П. А., Нгуен, К. Х., Гарридо, А. Л., Гиллеспи, Э. Р., и Харли, С. Б. Р. (2020). Повторное заселение микроглии способствует восстановлению мозга зависимым от IL-6 образом. Cell 180, 833.e–846.e. doi: 10.1016/j.cell.2020.02.013
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Винум Дж. Ю., Поулсен С. А. и Супуран С. Т. (2009 г.). Терапевтическое применение гликозидных ингибиторов карбоангидразы. Мед. Рез. 29, 419–435. doi: 10.1002/med.20141
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Син, К. , Хаякава, К., Лок, Дж., Араи, К., и Ло, Э. Х. (2012). Травма и восстановление в сосудисто-нервном блоке. Нейрол. Рез. 34, 325–330. doi: 10.1179/1743132812Y.0000000019
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ян К., Хокинс К. Э., Доре С. и Канделарио-Джалил Э. (2019 г.). Нейровоспалительные механизмы повреждения гематоэнцефалического барьера при ишемическом инсульте. утра. Дж. Физиол. Клетка. Физиол. 316, C135–C153. doi: 10.1152/ajpcell.00136.2018
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ян, Ю., и Розенберг, Г. А. (2015). Матриксные металлопротеиназы как терапевтические мишени при инсульте. Мозг Res. 1623, 30–38. doi: 10.1016/j.brainres.2015.04.024
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Яо, Ю. (2019). Базальная мембрана и инсульт. Дж. Цереб. Кровоток Метаб. 39, 3–19. doi: 10.1177/0271678X18801467
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Заманова С.