Технические характеристики двигателей для Фольксваген Транспортер Т5 GP модели 2010 года.
Технические характеристики двигателей для Фольксваген Транспортер Т5 GP модели 2010 года.2- литровый дизельный двигатель с системой впрыска Common-Rail для модели T5 2010 создан на базе двигателя TDI объемом 2,0 л модели Tiguan/Passat.
В T5 2010 используется 2-е поколение этих двигателей (EA 189).
Для T5 2010 предлагается четыре варианта мощности:
62 кВт (84 л.с.)
75 кВт (102 л.с.)
103 кВт (140 л.с.)
132
кВт (180 л.с.)
В дальнейшем этот двигатель будет также устанавливаться на модели Amarok
и
Crafter.
Уходят в
историю
5-цилиндровые 2,5 TDI
4-цилиндровые 1,9 TDI
Курс на внедрение
Common Rail вместо насос-форсунок.
Буквенное
обозначение двигателя CAAA
Рабочий объем 1968 см³
Количество цилиндров
4
Количество клапанов на цилиндр 4
Диаметр цилиндра 81,0 мм
Коэффициент сжатия 16,5 : 1
Макс.
 мощность 62 кВт при 3500
об/мин
мощность 62 кВт при 3500
об/минМакс. крутящий момент 220 Нм при
1250–2500 об/мин
Блок управления двигателем EDC 17CP 20
Турбокомпрессор с изменяемой геметрией
Рециркуляция ОГ да
Сажевый фильтр да (EU5)
нет (EU4)
Фольксваген Транспортер Т5 2010 Технические характеристики двигателей.
Буквенное
обозначение двигателя CAAB
Рабочий объем 1968 см³
Количество цилиндров
4
Количество клапанов на цилиндр 4
Диаметр цилиндра 81,0 мм
Ход поршня
95,5 мм
Коэффициент сжатия 16,5 : 1
Макс. мощность 75 кВт при 3500
об/мин
Макс. крутящий момент 250 Нм при
1500–2500 об/мин
Блок
управления двигателем EDC 17CP 20
Турбокомпрессор с изменяемой
геметрией
Рециркуляция ОГ да
Сажевый фильтр да (EU5)
нет
(EU4)
Буквенное
обозначение двигателя CAAC, CCHA
Количество цилиндров 4
Количество клапанов на цилиндр 4
Диаметр цилиндра 81,0 мм
Ход поршня 95,5 мм
Коэффициент сжатия 16,5 : 1
Макс.
 мощность 103
кВт при 3500 об/мин
мощность 103
кВт при 3500 об/минМакс. крутящий момент 340 Нм при
1750–2500 об/мин
Блок управления двигателем EDC 17CP 20
Турбокомпрессор с изменяемой геметрией
Рециркуляция ОГ да
Сажевый фильтр да (EU5)
нет (EU4)
Двигатель CCHA оснащен модулем балансирного вала (AGW)
Двигатель 2,0
л TDI с буквенным обозначением CAAC используется в различных
версиях модели
Transporter.
Двигатель 2,0 л TDI с буквенным обозначением CCHA предназначен
для моделей
Caravelle, Multivan и California (Comfortline). Этот двигатель
оснащен модулем
балансирного вала (AGW).
Буквенное
обозначение двигателя CFCA
Рабочий объем 1968 см³
Количество цилиндров
4
Количество клапанов на цилиндр 4
Диаметр цилиндра 81,0 мм
Ход поршня
95,5 мм
Коэффициент сжатия 16,5 : 1
Макс. мощность 132 кВт при 4000
об/мин
Макс. крутящий момент 400 Нм при
1500–2250 об/мин
Блок
управления двигателем EDC 17CP 20
Турбокомпрессор: Модуль двойного
наддува
Рециркуляция ОГ да
Сажевый фильтр да (EU5)
нет
(EU4)
Двигатели T5 2010 .![]() Технические
характеристики.
Технические
характеристики.
Общее представление автомобиля Volkswagen Transporter T5 модели 2010 года.
Volkswagen Transporter T5 2010 , автомобиль внутри.
VW T5 2010 Механика двигателя Common-Rail с турбонаддувом
продолжениеБезопасность VW T5 2010 .
Электрооборудование VW T5 2010 .
Volkswagen Transporter T5 2010 , ходовая часть.
Двигатели T5 2010 .Технические характеристики.
Двигатель 2,0 л Common-Rail с турбонаддувом
VW T5 2010 Механика двигателя Common-Rail с турбонаддувом
Двигатель 2,0 л TDI BiTurbo с двойным наддувом
Механика двигателя TDI BiTurbo с двойным наддувом
Турбокомпрессор / Двигатель 2,0 л TDI BiTurbo с двойным наддувом
Трансмиссия VW
T5 2010 . Кинематическая схема коробки передач
Фольксваген Т5 2010
Кинематическая схема коробки передач
Фольксваген Т5 2010
Трансмиссия VW T5 2010 . Монтажные схемы КПП
Трансмиссия VW T5 2010 . Механизм переключения передач
Трансмиссия VW T5 2010 . Двойное сцепление
Трансмиссия VW T5 2010 . Мехатроник
- Обои для рабочего стола
- Авторские фотографии
- Volkswagen Transporter / Видео / Crash тесты / Ролики
- Ремонт и обслуживание
- Электрооборудование
- Книги по ремонту и обслуживанию
- Официальные дилеры и автосалоны
- Volkswagen Transporter T5 GP 2010
- Технические характеристики
- Статьи и отзывы владельцев
Фотогалерея
Партнеры
Russian Team
VW-Transporter. ru ®
ru ®
All rights reserved © 2008
Двигатель и
трансмиссия .
Обсуждение технических вопросов : система
смазки , система охлаждения , система выпуска отработавших газов , системы
впрыска топлива .
Электрооборудование .
Обсуждение
технических вопросов : датчики , реле , предохранители , АКБ ,генератор ,
система освещения , кондиционер , отопитель .
Шасси
.
Обсуждение технических вопросов : передний мост , задний
мост , рулевое управление , тормозная система , колеса и шины .
Кузов
.
Обсуждаем наружную и внутреннюю часть кузова , замки ,
двери , регулировки внешних и внутренних элементов , как снять и как поставить и
другие вопросы .
Тюнинг
.
Здесь обсуждаем внешний и внутренний нестандартный тюнинг :
тонировка , бампера , сигнализации , ксенон , салон , аэрография , шины и диски
, техническое оборудование , электроника , музыка , навигационное оборудование .
Автосервисы
, авто ателье и магазины авто запчастей .
Обсуждаем и
оставляем отзывы о качестве обслуживания : где лучше ремонтироваться , покупать
запчасти , проводить переоборудование или делать тюнинг автомобиля Фольксваген
Транспортер .
Полезные
ссылки .
Ссылки на полезные странички , электронные книги по
ремонту и обслуживанию автомобилей VW Transporter , каталоги автозапчастей ,
автомобильные журналы , и другие файлы для владельцев Фольксваген Транспортер .
Форум владельцев VOLKSWAGEN TRANSPORTER T5 RusTeam .
Общаемся и
советуемся , делимся личным опытом эксплуатации и обслуживания автомобилей
Фольксваген Транспортер , помогаем другим участника форума разобраться в
возникших сложностях и проблемах .
Объявления
ООО «Рустурбоком»
Ремонт турбин, ТНВД , а также
оптовая и розничная продажа турбокомпрессоров ведущих мировых производителей для
легковых , грузовых автомобилей и автобусов.
Главная
Контакты
Архивный форум
Характеристики Т5 — Электронный марочник российских сталей
Switch to English
:- Стандарты
- Химсостав
- Свойства
- Другие наименования
- Применение
- Аналоги
| Si (Кремний) | |
| Mn (Марганец) | |
| P (Фосфор) | |
| S (Сера) | |
| Cr (Хром) | |
| Ni (Никель) | |
| Al (Алюминий) | |
| Cu (Медь) | |
| N (Азот) | |
| Fe (Железо) | |
| Ca (Кальций) |
Примечание:
лавочный анализ
| ГОСТ Р 52204-04 |
Для получения информации о зарубежных аналогах данной стали воспользуйтесь нашим бесплатным Сервисом запросов.
- Прайс-листы
- Цены LME
- Фосс Металл (152 поз.)
- ООО ПКП «Теплообменные трубы» (582 поз.)
- Снабтехмет-Новосибирск (20000 поз.)
- НПК «Специальная металлургия-Новосибирск» (20000 поз.)
- СТМ-Восток (20000 поз.)
- НПК «Специальная металлургия-Владивосток» (3240 поз.)
Поиск по прайс-листам
Все компании
| Цены LME | |||
| Cash seller & settlement | |||
| Cobalt | 56570,60 | (0,00) | |
| Steel Scrap | 364,00 | (1,00) | |
| Steel Rebar | 680,00 | (7,00) | |
| NASAAC | 2495,00 | (-5,00) | |
| Aluminium | 2162,00 | (-35,00) | |
| Copper | 7560,00 | (18,00) | |
| Zinc | 2968,00 | (38,00) | |
| Nickel | 21107,00 | (-1241,00) | |
| Lead | 1908,00 | (32,00) | |
| Aluminium Alloy | 1760,00 | (50,00) | |
| Tin | 20634,00 | (111,00) | |
Марочник сталей Т5 РС E40Z35 77НМДП
Мониторинг цен
Центральный регион, Москва
22 сентября 2022 года
| Сортамент | Средняя цена | Индекс |
|---|---|---|
Арм. А500С ф10 А500С ф10 | 50,893.8 | -0.2 % |
| Арм.А500С ф12 | 49,250.0 | -0.1 % |
| Арм. В500С ф8 | 51,390.0 | -0.1 % |
| Проволока вр ф4-5 | 47,060.0 | -0.1 % |
| Проволока ок ф1,2 | 85,400.0 | -0.8 % |
| Катанка ф6,5 | 49,260. 0 0 | -0.2 % |
| Лист г/к 4 | 55,428.6 | 1.7 % |
| Лист х/к 0,8-1 | 61,716.7 | 1.9 % |
| Лист оц. 0,55 | 78,380.0 | 7.4 % |
| Труба ВГП 20х2,8 | 64,566.7 | 0.9 % |
| Труба ВГП 32х3,2 | 57,883.3 | 1. 9 % 9 % |
| Труба э/с 89х3,5 | 52,766.7 | 2.8 % |
| Труба э/с 102х3,5 | 53,600.0 | 2.7 % |
| Уголок р/п 63х5-6 | 61,100.0 | 0.0 % |
| Швеллер 10 | 70,214.3 | -2.2 % |
| Швеллер 12 | 73,166.7 | -0.9 % |
| Балка 30Б1 | 65,300. 0 0 | 0.0 % |
| Балка 30Ш1 | 62,900.0 | 0.0 % |
| Динамика: | 0.9 % | |
Опрос МСС
Что вы ждете от осени т.г.?
(проводился с 30-08 по 18-09-2022)- Стагнацию и падение цен на стальной прокат 69 (34,16%)
- Неопределенность продолжится 45 (22,28%)
- Очередных потрясений на рынке металлов и не только 31 (15,35%)
- Роста спроса и цен на стальную продукцию 29 (14,36%)
- Стабильных цен и устойчивого спроса со стороны стройкомплекса 28 (13,86%)
- Всего голосов: 202
- Завершенные опросы
Металлургическая мозаика:
В 2017 г..jpg) профессиональный борец Джон Ферарро установил необычное достижение:
профессиональный борец Джон Ферарро установил необычное достижение:
за три минуты он забил головой в доску 38 гвоздей. Для большинства людей это может оказаться смертельным, но у Джона Ферарро кости черепа гораздо толще.
Читать далее
KAMAZ-65801-68(Т5)
Заказать
Стандарт: Евро 5. Колёсная формула: 8х4
Технические характеристики
- Весовые параметры и нагрузки
Грузоподъемность автомобиля, кг 32435 Полная масса а/м, кг 50000 нагрузка на заднюю тележку, кг 32000 нагрузка на первую и вторую оси, кг 18000 Снаряженная масса, кг 17565 нагрузка на заднюю тележку, кг 8045 нагрузка на первую и вторую оси, кг 9520 - Двигатель
Макс.  полезный крутящий момент, Нм (кгсм)
полезный крутящий момент, Нм (кгсм)2100 (214) при частоте вращения коленчатого вала, об/мин 1100 Максимальная полезная мощность, кВт (л.с.) 315 (428) при частоте вращения коленчатого вала, об/мин 1900 Рабочий объем, л 11,97 Расположение и число цилиндров рядное, 6 Тип двигателя дизельный с турбонаддувом, с промежуточным охлаждением наддувочного воздуха - Коробка передач
Тип механическая, 16-тиступенчатая Управление механическое, дистанционное - Габаритные размеры, мм
Длина, мм 9280 Ширина, мм 2550 Высота, мм 3450 - Главная передача
Передаточное отношение 5,262 Тип двойная, с колесными редукторами - Кабина
Исполнение без спального места Подвеска 4-х точечная пружинная Тип кабины расположенная над двигателем - Колеса и шины
Размер обода 8,5-24 Тип колес дисковые Тип шин пневматические, камерные Шины 12. 00 R24 - Самосвальная платформа
Исполнение обогрев выхлопными газами, полог с механизмом скручивания, дополнительная лестница Материал платформы днище-8 мм, борта-5 мм; износостойкая сталь, твердость 450 HB Объем платформы, куб. м 20 Угол подъема платформы, град 50 - Система питания
Вместимость бака с нейтрализующей жидкостью, л 70 Вместимость топливного бака, л 350 - Сцепление
Привод гидравлический с пневмоусилителем Тип однодисковое, диафрагменное мод. MFZ 430 - Тормоза
Привод пневматический Размеры диаметр барабана, мм 420 Ширина тормозных накладок, мм 180 - Характеристики а/м полной массой
Внешний габаритный радиус поворота, м 11,6 Максимальная скорость, не менее, км/ч 90 Угол преодолеваемого подъема, не менее, % (град) 30 (18) - Электрооборудование
Аккумуляторы, В/А·ч 2х12/190 Напряжение, B 24
 полезный крутящий момент, Нм (кгсм)
полезный крутящий момент, Нм (кгсм) 00 R24
00 R24 MFZ 430
MFZ 430Вся представленная на сайте информация, касающаяся технических характеристик, наличия на складе, стоимости товаров,
носит информационный характер и ни при каких условиях не является публичной офертой, определяемой положениями Статьи 437(2) Гражданского кодекса РФ.
Поломоечная машина Tennant Т5 — характеристики, подробное описание
Данная модель снята с производства. Предлагаем улучшенный аналог Tennant T500.
- Наличие на складе
- Покупка в лизинг
Основные характеристики:
Привод
АКБ
Ширина уборки
80 см
Производительность
2660 м2
Полные характеристики
Официальные поставщики
Доставка от 1 дня
Обученный персонал
Точки ремонта по всей РФ
Качественные запчасти
Общее описание Технические характеристики Доставка и оплата
Общее описание
Инструкции и брошюры Tennant T5
- Брошюра Tennant T5
- Руководство оператора Tennant T5
Достигните высочайшего уровня чистоты, гигиены и безопасности с поломоечной машиной Tennant T5
- Берегите ресурсы! Чистите без химикатов с новой технологией ec-h3O™ — электрически активированная вода превращает воду в мощное чистящее средство
- Берегите ресурсы! Чистите, используя на 90% меньше чистящего средства, и уменьшите размеры упаковки для химикатов на 80% с технологией FaST®
- Берегите ресурсы! Уменьшите потребление воды на 90% и сократите неэффективное использование на 70% с технологией FaST®
- Поломоечная машина tennant T5 уменьшает количество бактерий и налета благодаря легко очищаемым и легко доступным гигиеничным бакам
- Низкий уровень шума в 67 дБ позволяет чистить в самых чувствительных к шуму местах
- Улучшенная безопасность — поломоечная машина Tennant T5 полностью удаляет моющее средство, даже при повороте на 180 градусов, благодаря уникально спроектированному скребку
Ключевые характеристики производительности и надежности машины Tennant T5:
- В машине Tennant Т5 установлены гигиеничные баки, уменьшающие количество плесени, бактерий и других загрязняющих веществ
- Работающая от аккумуляторов поломоечная машина Tennant T5 практически не издает шума
- Параболическая система скребка обладает оптимальным углом атаки, что позволяет полностью удалять остатки моющего средства
- Технология FaST® использует до 70% меньше моющего средства во время чистки
- Технология ec-h3O™ обеспечивает великолепный результат чистки без использования химикатов
Выберите лучшую технологию чистки для вашей среды
- Новая технология ec-h3O™ — электрически активированная вода сокращает негативное влияние на окружающую среду: изготовление упаковки, транспортировку, использование и переработку химикатов
- Технология FaST® — технология чистки пеной. Используя инновационную систему чистки пеной, поломоечные машины с FaST увеличивают производительность до 30%, уменьшают потребление воды до 70% и исключают необходимость контакта оператора с химикатами.
 Используя инновационную систему чистки пеной, поломоечные машины с FaST увеличивают производительность до 30%, уменьшают потребление воды до 70% и исключают необходимость контакта оператора с химикатами.
Используя инновационную систему чистки пеной, поломоечные машины с FaST увеличивают производительность до 30%, уменьшают потребление воды до 70% и исключают необходимость контакта оператора с химикатами.Tennant T5 – среднеразмерная производительная машина, для работы которой используется технология FaST®, снижающая объем химических моющих средств на 80%. Кроме того, значительно снижено потребление воды, улучшены показатели производительности, машина оборудована удобными гигиеническими баками.
Техника может использоваться для жилых помещений, офисов, торговых и развлекательных центров, салонов. В качестве источника питания установлена батарея на 24 В, общая мощность техники – 1200 Вт.
Особенности и преимущества модели Т5
Моющая машина T5 среднеразмерного класса отличается такими преимуществами:
- производитель – до 1000-2500 кв.м/час;
- ширина дорожки очистки – 60-70см;
- скорость уборки (в зависимости от выбранного режима) – 2925-3600 кв. м/час;
- низкий уровень шума – 67 ДБа;
- простая замена щеток;
- наличие технологии FaST® для экономного расхода воды и моющих средств;
- автономность, благодаря установленному аккумулятору;
- возможность использования модели с технологий ec-h3O.
 м/час;
м/час;Модель является самой мощной в среднеразмерном классе, она востребована для уборки офисов, развлекательных центров, жилых помещений, салонов и магазинов. Удобные щетки обеспечивают уборку мусора в труднодоступных места, машина работает от батареи, что обеспечивает ее автономность и мобильность.
Технические характеристики
Дисковая | Цилиндрическая | ||
|---|---|---|---|
Размеры поломоечной машины Tennant T5 | |||
| Ширина убираемой поверхности | см | 60-70-80 | 65-80 |
| Ширина уборочного узла | см | 91-105-119 | 105-119 |
| Вес машины | кг | 143-152-158 | 162-166 |
| Размеры (длина x ширина x высота) | см | 135×65/75/85×112 | 135×73/88×112 |
Моющая система поломоечной машины Tennant T5 | |||
| Скорость уборки теоретическая (макс) | кв м / час | 2700-3150-3600 | 2925-3600 |
| Скорость уборки практическая (макс) | кв м / час | 1375-1625-1875 | 1500-1875 |
| Количество щеток, падов | шт. | 2 | 2 |
| Диаметр дисковых щетов, падов | см | 30-35-40 | — |
| Длина x диаметр цилиндрических щёток | см | — | 66/81×15 |
Прочие параметры поломоечной машины Tennant T5 | |||
| Воздушный поток | л/сек | 33 | 33 |
| Подъем воды | кПа | 15,5 | 15,5 |
| Практическая емкость бака для воды | л | 85 | 85 |
| Практическая емкость бака для грязной воды | л | 102 | 102 |
Питание поломоечной машины Tennant T5 | |||
| Источник питания | батарея 24 В | батарея 24 В | |
| Полная потребляемая мощность (стандарт) | Вт | 1200 | 1200 |
| Класс защиты | I/II/III | III | III |
| Уровень шума (DIN45635) | Дб (А) | 67 | 68 |
Варианты поломоечной машины Tennant T5 | |||
| ec-h3O | да/ нет | да | да |
| FaST | да/ нет | да | да |
| Цилиндрический / дисковый | дисковый | цилиндрический | |
Транспортные характеристики поломоечной машины Tennant T5 | |||
| Транспортная скорость | км/час | 4,5 | 4,5 |
| Рабочая скорость | км/час | 2,5 | 2,5 |
| Максимальный угол подъема | % | 8 | 8 |
| Минимальное расстояние для разворота | см | 135-150-163 | 157-164 |
Доставка и оплата
- Мы можем организовать доставку нашей техники до любой точки России — от Калининграда до Петропавловска-Камчатского.
- Обычно доставка осуществляется транспортной компанией «Деловые линини», но если вам удобнее работать с другой компанией — никаких проблем, мы работаем со всеми операторами.
- Все отправляемые нами грузы аккуратно упаковываются, и страхуются.

ЗАКАЖИТЕ БЕСПЛАТНЫЙ ТЕСТ-ДРАЙВ
и получите персональную скидку 5%
Ваше имя Телефон s
Согласен на обработку персональных данных
похожие модели
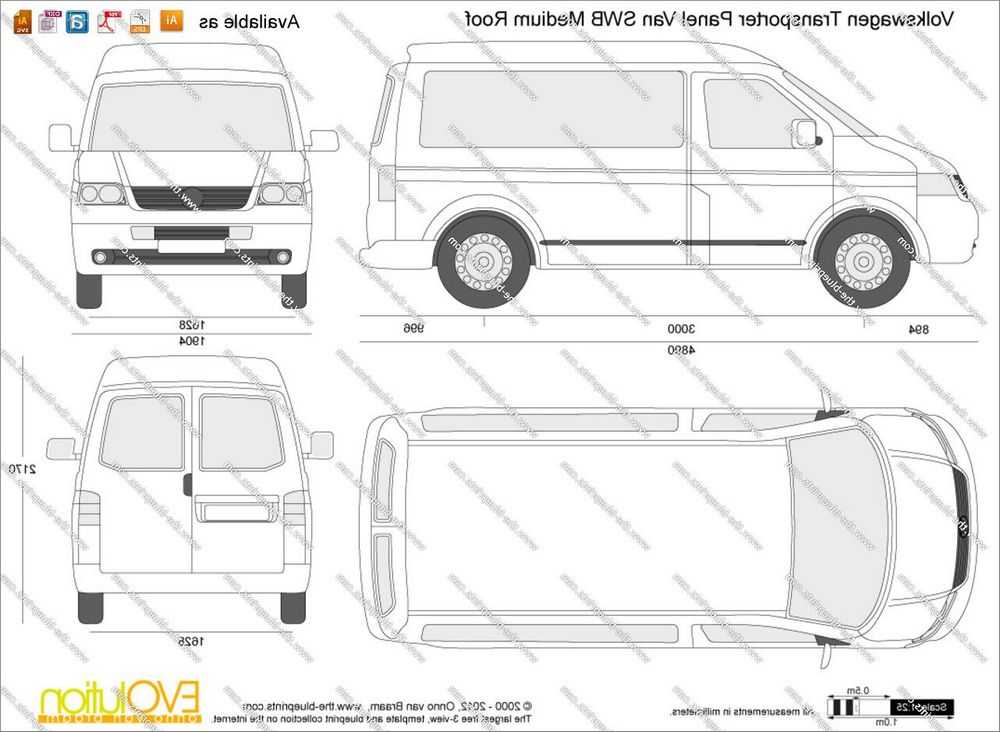
Технические характеристики Volkswagen Transporter furgon с 2010 года: подробно — Бибипедия
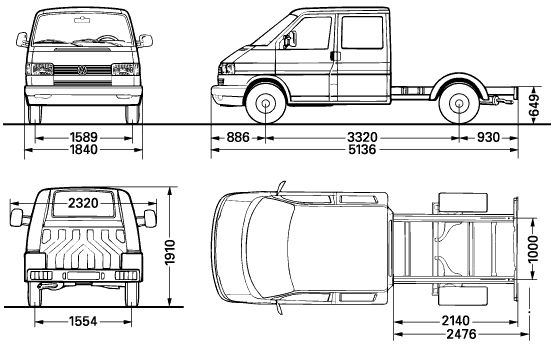
Технические характеристики Volkswagen Transporter 2010, 2011, 2012, 2013: мощность, расход топлива на 100 км, вес (масса), дорожный просвет (клиренс), радиус разворота, тип трансмиссии и тормозов, размеров кузова и шин
Характеристики двигателя
| Модификации | Объём двигателя, см3 | Мощность, кВт (л.с.)/об | Цилиндры | Крутящий момент, Нм/(об/мин) | Тип топливной системы | Тип топлива |
|---|---|---|---|---|---|---|
2. 0 BiTDI DSG 4Motion L1h2 0 BiTDI DSG 4Motion L1h2 | 1968 | (180)/4000 | Рядный — 4 | (400)/1500-2000 | Непосредственный впрыск | Дизель |
| 2.0 BiTDI DSG 4Motion L1h3 | 1968 | (180)/4000 | Рядный — 4 | (400)/1500-2000 | Непосредственный впрыск | Дизель |
| 2.0 BiTDI DSG 4Motion L2h2 | 1968 | (180)/4000 | Рядный — 4 | (400)/1500-2000 | Непосредственный впрыск | Дизель |
2. 0 BiTDI DSG 4Motion L2h3 0 BiTDI DSG 4Motion L2h3 | 1968 | (180)/4000 | Рядный — 4 | (400)/1500-2000 | Непосредственный впрыск | Дизель |
| 2.0 BiTDI DSG L1h2 | 1968 | (180)/4000 | Рядный — 4 | (400)/1500-2000 | Непосредственный впрыск | Дизель |
| 2.0 BiTDI DSG L1h3 | 1968 | (180)/4000 | Рядный — 4 | (400)/1500-2000 | Непосредственный впрыск | Дизель |
| 2.0 BiTDI DSG L2h2 | 1968 | (180)/4000 | Рядный — 4 | (400)/1500-2000 | Непосредственный впрыск | Дизель |
| 2. | 1968 | (180)/4000 | Рядный — 4 | (400)/1500-2000 | Непосредственный впрыск | Дизель |
| 2.0 BiTDI DSG L2h4 | 1968 | (180)/4000 | Рядный — 4 | (400)/1500-2000 | Непосредственный впрыск | Дизель |
| 2.0 BiTDI MT 4Motion L1h2 | 1968 | (180)/4000 | Рядный — 4 | (400)/1500-2000 | Непосредственный впрыск | Дизель |
| 2.0 BiTDI MT 4Motion L1h3 | 1968 | (180)/4000 | Рядный — 4 | (400)/1500-2000 | Непосредственный впрыск | Дизель |
2. 0 BiTDI MT 4Motion L2h2 0 BiTDI MT 4Motion L2h2 | 1968 | (180)/4000 | Рядный — 4 | (400)/1500-2000 | Непосредственный впрыск | Дизель |
| 2.0 BiTDI MT 4Motion L2h3 | 1968 | (180)/4000 | Рядный — 4 | (400)/1500-2000 | Непосредственный впрыск | Дизель |
| 2.0 BiTDI MT 4Motion L2h4 | 1968 | (180)/4000 | Рядный — 4 | (400)/1500-2000 | Непосредственный впрыск | Дизель |
| 2. | 1968 | (180)/4000 | Рядный — 4 | (400)/1500-2000 | Непосредственный впрыск | Дизель |
| 2.0 BiTDI MT L1h3 | 1968 | (180)/4000 | Рядный — 4 | (400)/1500-2000 | Непосредственный впрыск | Дизель |
| 2.0 BiTDI MT L2h2 | 1968 | (180)/4000 | Рядный — 4 | (400)/1500-2000 | Непосредственный впрыск | Дизель |
| 2.0 BiTDI MT L2h3 | 1968 | (180)/4000 | Рядный — 4 | (400)/1500-2000 | Непосредственный впрыск | Дизель |
| 2. | 1968 | (180)/4000 | Рядный — 4 | (400)/1500-2000 | Непосредственный впрыск | Дизель |
| 2.0 MT L1h2 | 1984 | (115)/5200 | Рядный — 4 | (170)/2700-4700 | Инжектор | Бензин |
| 2.0 MT L1h3 | 1984 | (115)/5200 | Рядный — 4 | (170)/2700-4700 | Инжектор | Бензин |
| 2.0 MT L2h2 | 1984 | (115)/5200 | Рядный — 4 | (170)/2700-4700 | Инжектор | Бензин |
| 2. | 1984 | (115)/5200 | Рядный — 4 | (170)/2700-4700 | Инжектор | Бензин |
| 2.0 MT L2h4 | 1984 | (115)/5200 | Рядный — 4 | (170)/2700-4700 | Инжектор | Бензин |
| 2.0 TDI DSG L1h2 | 1968 | (140)/3500 | Рядный — 4 | (340)/1750-2500 | Непосредственный впрыск | Дизель |
| 2.0 TDI DSG L1h3 | 1968 | (140)/3500 | Рядный — 4 | (340)/1750-2500 | Непосредственный впрыск | Дизель |
2. 0 TDI DSG L2h2 0 TDI DSG L2h2 | 1968 | (140)/3500 | Рядный — 4 | (340)/1750-2500 | Непосредственный впрыск | Дизель |
| 2.0 TDI DSG L2h3 | 1968 | (140)/3500 | Рядный — 4 | (340)/1750-2500 | Непосредственный впрыск | Дизель |
| 2.0 TDI DSG L2h4 | 1968 | (140)/3500 | Рядный — 4 | (340)/1750-2500 | Непосредственный впрыск | Дизель |
| 2.0 TDI MT 4Motion L1h2 | 1968 | (140)/3500 | Рядный — 4 | (340)/1750-2500 | Непосредственный впрыск | Дизель |
2. 0 TDI MT 4Motion L1h3 0 TDI MT 4Motion L1h3 | 1968 | (140)/3500 | Рядный — 4 | (340)/1750-2500 | Непосредственный впрыск | Дизель |
| 2.0 TDI MT 4Motion L2h2 | 1968 | (140)/3500 | Рядный — 4 | (340)/1750-2500 | Непосредственный впрыск | Дизель |
| 2.0 TDI MT 4Motion L2h3 | 1968 | (140)/3500 | Рядный — 4 | (340)/1750-2500 | Непосредственный впрыск | Дизель |
2. 0 TDI MT 4Motion L2h4 0 TDI MT 4Motion L2h4 | 1968 | (140)/3500 | Рядный — 4 | (340)/1750-2500 | Непосредственный впрыск | Дизель |
| 2.0 TDI MT L1h2 | 1968 | (140)/3500 | Рядный — 4 | (340)/1750-2500 | Непосредственный впрыск | Дизель |
| 2.0 TDI MT L1h3 | 1968 | (140)/3500 | Рядный — 4 | (340)/1750-2500 | Непосредственный впрыск | Дизель |
| 2.0 TDI MT L2h2 | 1968 | (140)/3500 | Рядный — 4 | (340)/1750-2500 | Непосредственный впрыск | Дизель |
2. 0 TDI MT L2h3 0 TDI MT L2h3 | 1968 | (140)/3500 | Рядный — 4 | (340)/1750-2500 | Непосредственный впрыск | Дизель |
| 2.0 TDI MT L2h4 | 1968 | (140)/3500 | Рядный — 4 | (340)/1750-2500 | Непосредственный впрыск | Дизель |
Привод и трансмиссия
| Модификации | Тип привода | Тип трансмиссии (базовая) | Тип трансмиссии (опционально) |
|---|---|---|---|
| 2.0 BiTDI DSG 4Motion L1h2 | 4×4 (Полный привод) | Автоматическая с двумя сцеплениями — 7 | |
2. 0 BiTDI DSG 4Motion L1h3 0 BiTDI DSG 4Motion L1h3 | 4×4 (Полный привод) | Автоматическая с двумя сцеплениями — 7 | |
| 2.0 BiTDI DSG 4Motion L2h2 | 4×4 (Полный привод) | Автоматическая с двумя сцеплениями — 7 | |
| 2.0 BiTDI DSG 4Motion L2h3 | 4×4 (Полный привод) | Автоматическая с двумя сцеплениями — 7 | |
| 2.0 BiTDI DSG L1h2 | Передний привод | Автоматическая с двумя сцеплениями — 7 | |
| 2.0 BiTDI DSG L1h3 | Передний привод | Автоматическая с двумя сцеплениями — 7 | |
2. 0 BiTDI DSG L2h2 0 BiTDI DSG L2h2 | Передний привод | Автоматическая с двумя сцеплениями — 7 | |
| 2.0 BiTDI DSG L2h3 | Передний привод | Автоматическая с двумя сцеплениями — 7 | |
| 2.0 BiTDI DSG L2h4 | Передний привод | Автоматическая с двумя сцеплениями — 7 | |
| 2.0 BiTDI MT 4Motion L1h2 | 4×4 (Полный привод) | 6-МКПП | |
| 2.0 BiTDI MT 4Motion L1h3 | 4×4 (Полный привод) | 6-МКПП | |
| 2.0 BiTDI MT 4Motion L2h2 | 4×4 (Полный привод) | 6-МКПП | |
2. 0 BiTDI MT 4Motion L2h3 0 BiTDI MT 4Motion L2h3 | 4×4 (Полный привод) | 6-МКПП | |
| 2.0 BiTDI MT 4Motion L2h4 | 4×4 (Полный привод) | 6-МКПП | |
| 2.0 BiTDI MT L1h2 | Передний привод | 6-МКПП | |
| 2.0 BiTDI MT L1h3 | Передний привод | 6-МКПП | |
| 2.0 BiTDI MT L2h2 | Передний привод | 6-МКПП | |
| 2.0 BiTDI MT L2h3 | Передний привод | 6-МКПП | |
2..jpg) 0 BiTDI MT L2h4 0 BiTDI MT L2h4 | Передний привод | 6-МКПП | |
| 2.0 MT L1h2 | Передний привод | 5-МКПП | |
| 2.0 MT L1h3 | Передний привод | 5-МКПП | |
| 2.0 MT L2h2 | Передний привод | 5-МКПП | |
| 2.0 MT L2h3 | Передний привод | 5-МКПП | |
| 2.0 MT L2h4 | Передний привод | 5-МКПП | |
2. 0 TDI DSG L1h2 0 TDI DSG L1h2 | Передний привод | Автоматическая с двумя сцеплениями — 7 | |
| 2.0 TDI DSG L1h3 | Передний привод | Автоматическая с двумя сцеплениями — 7 | |
| 2.0 TDI DSG L2h2 | Передний привод | Автоматическая с двумя сцеплениями — 7 | |
| 2.0 TDI DSG L2h3 | Передний привод | Автоматическая с двумя сцеплениями — 7 | |
| 2.0 TDI DSG L2h4 | Передний привод | Автоматическая с двумя сцеплениями — 7 | |
2. 0 TDI MT 4Motion L1h2 0 TDI MT 4Motion L1h2 | 4×4 (Полный привод) | 6-МКПП | |
| 2.0 TDI MT 4Motion L1h3 | 4×4 (Полный привод) | 6-МКПП | |
| 2.0 TDI MT 4Motion L2h2 | 4×4 (Полный привод) | 6-МКПП | |
| 2.0 TDI MT 4Motion L2h3 | 4×4 (Полный привод) | 6-МКПП | |
| 2.0 TDI MT 4Motion L2h4 | 4×4 (Полный привод) | 6-МКПП | |
| 2.0 TDI MT L1h2 | Передний привод | 6-МКПП | |
2. 0 TDI MT L1h3 0 TDI MT L1h3 | Передний привод | 6-МКПП | |
| 2.0 TDI MT L2h2 | Передний привод | 6-МКПП | |
| 2.0 TDI MT L2h3 | Передний привод | 6-МКПП | |
| 2.0 TDI MT L2h4 | Передний привод | 6-МКПП |
Тип подвески и радиус разворота
| Модификации | Передняя подвеска | Задняя подвеска | Диаметр разворота,м |
|---|---|---|---|
| 2.0 BiTDI DSG 4Motion L1h2 | Независимая — многорычажная | Независимая — многорычажная | — |
| 2. | Независимая — многорычажная | Независимая — многорычажная | — |
| 2.0 BiTDI DSG 4Motion L2h2 | Независимая — многорычажная | Независимая — многорычажная | — |
| 2.0 BiTDI DSG 4Motion L2h3 | Независимая — многорычажная | Независимая — многорычажная | — |
| 2.0 BiTDI DSG L1h2 | Независимая — McPherson | Полузависимая — торсионная балка | 9.96 |
| 2.0 BiTDI DSG L1h3 | Независимая — McPherson | Полузависимая — торсионная балка | 9. 96 96 |
| 2.0 BiTDI DSG L2h2 | Независимая — McPherson | Полузависимая — торсионная балка | 9.96 |
| 2.0 BiTDI DSG L2h3 | Независимая — McPherson | Полузависимая — торсионная балка | 9.96 |
| 2.0 BiTDI DSG L2h4 | Независимая — McPherson | Полузависимая — торсионная балка | 10.6 |
| 2.0 BiTDI MT 4Motion L1h2 | Независимая — McPherson | Полузависимая — торсионная балка | 10.6 |
| 2.0 BiTDI MT 4Motion L1h3 | Независимая — McPherson | Полузависимая — торсионная балка | 10. 4 4 |
| 2.0 BiTDI MT 4Motion L2h2 | Независимая — McPherson | Полузависимая — торсионная балка | 10.4 |
| 2.0 BiTDI MT 4Motion L2h3 | Независимая — McPherson | Полузависимая — торсионная балка | 10.4 |
| 2.0 BiTDI MT 4Motion L2h4 | Независимая — McPherson | Полузависимая — торсионная балка | 10.4 |
| 2.0 BiTDI MT L1h2 | Независимая — McPherson | Полузависимая — торсионная балка | 10.6 |
| 2.0 BiTDI MT L1h3 | Независимая — McPherson | Полузависимая | — |
2. 0 BiTDI MT L2h2 0 BiTDI MT L2h2 | Независимая — McPherson | Полузависимая | — |
| 2.0 BiTDI MT L2h3 | Независимая — McPherson | Полузависимая | — |
| 2.0 BiTDI MT L2h4 | Независимая — McPherson | Полузависимая — торсионная балка | — |
| 2.0 MT L1h2 | Независимая — McPherson | Полузависимая — торсионная балка | 10.7 |
| 2.0 MT L1h3 | Независимая — McPherson | Полузависимая — торсионная балка | — |
| 2. | Независимая — McPherson | Полузависимая — торсионная балка | 10.7 |
| 2.0 MT L2h3 | Независимая — McPherson | Полузависимая — торсионная балка | 10.7 |
| 2.0 MT L2h4 | Независимая — McPherson | Полузависимая — торсионная балка | — |
| 2.0 TDI DSG L1h2 | Независимая — McPherson | Полузависимая — торсионная балка | 10.7 |
| 2.0 TDI DSG L1h3 | Независимая — McPherson | Полузависимая — торсионная балка | 10. |
| 2.0 TDI DSG L2h2 | Независимая — McPherson | Полузависимая — торсионная балка | 10.7 |
| 2.0 TDI DSG L2h3 | Независимая — McPherson | Полузависимая — торсионная балка | 10.7 |
| 2.0 TDI DSG L2h4 | Независимая — McPherson | Независимая — многорычажная | 10.6 |
| 2.0 TDI MT 4Motion L1h2 | Независимая — McPherson | Независимая — многорычажная | 10.6 |
| 2.0 TDI MT 4Motion L1h3 | Независимая — McPherson | Независимая — многорычажная | 10. 6 6 |
| 2.0 TDI MT 4Motion L2h2 | Независимая — McPherson | Независимая — многорычажная | 10.6 |
| 2.0 TDI MT 4Motion L2h3 | Независимая — McPherson | Независимая — многорычажная | 10.6 |
| 2.0 TDI MT 4Motion L2h4 | Независимая — McPherson | Независимая — многорычажная | — |
| 2.0 TDI MT L1h2 | Независимая — McPherson | Независимая — многорычажная | — |
| 2.0 TDI MT L1h3 | Независимая — McPherson | Независимая — многорычажная | — |
2. 0 TDI MT L2h2 0 TDI MT L2h2 | Независимая — McPherson | Независимая — многорычажная | — |
| 2.0 TDI MT L2h3 | Независимая — McPherson | Независимая — многорычажная | — |
| 2.0 TDI MT L2h4 | Независимая — McPherson | Независимая — многорычажная | — |
Тормозная система и усилитель руля
| Модификации | Тип передних тормозов | Тип задних тормозов | Усилитель руля |
|---|---|---|---|
| 2.0 BiTDI DSG 4Motion L1h2 | Дисковые вентилируемые | Дисковые | Гидравлический |
| 2. | Дисковые вентилируемые | Дисковые | Гидравлический |
| 2.0 BiTDI DSG 4Motion L2h2 | Дисковые вентилируемые | Дисковые | Гидравлический |
| 2.0 BiTDI DSG 4Motion L2h3 | Дисковые вентилируемые | Дисковые | Гидравлический |
| 2.0 BiTDI DSG L1h2 | Дисковые вентилируемые | Дисковые | Гидравлический |
| 2.0 BiTDI DSG L1h3 | Дисковые вентилируемые | Дисковые | Гидравлический |
2. 0 BiTDI DSG L2h2 0 BiTDI DSG L2h2 | Дисковые вентилируемые | Дисковые | Гидравлический |
| 2.0 BiTDI DSG L2h3 | Дисковые вентилируемые | Дисковые | Гидравлический |
| 2.0 BiTDI DSG L2h4 | Дисковые вентилируемые | Дисковые | Гидравлический |
| 2.0 BiTDI MT 4Motion L1h2 | Дисковые вентилируемые | Дисковые | Гидравлический |
| 2.0 BiTDI MT 4Motion L1h3 | Дисковые вентилируемые | Дисковые | Гидравлический |
| 2. | Дисковые вентилируемые | Дисковые | Гидравлический |
| 2.0 BiTDI MT 4Motion L2h3 | Дисковые вентилируемые | Дисковые | Гидравлический |
| 2.0 BiTDI MT 4Motion L2h4 | Дисковые вентилируемые | Дисковые | Гидравлический |
| 2.0 BiTDI MT L1h2 | Дисковые вентилируемые | Дисковые | Гидравлический |
| 2.0 BiTDI MT L1h3 | Дисковые вентилируемые | Дисковые | Гидравлический |
2. 0 BiTDI MT L2h2 0 BiTDI MT L2h2 | Дисковые вентилируемые | Дисковые | Гидравлический |
| 2.0 BiTDI MT L2h3 | Дисковые вентилируемые | Дисковые | Гидравлический |
| 2.0 BiTDI MT L2h4 | Дисковые вентилируемые | Дисковые | Гидравлический |
| 2.0 MT L1h2 | Дисковые вентилируемые | Дисковые | Гидравлический |
| 2.0 MT L1h3 | Дисковые вентилируемые | Дисковые | Гидравлический |
| 2.0 MT L2h2 | Дисковые вентилируемые | Дисковые | Гидравлический |
2. 0 MT L2h3 0 MT L2h3 | Дисковые вентилируемые | Дисковые | Гидравлический |
| 2.0 MT L2h4 | Дисковые вентилируемые | Дисковые | Гидравлический |
| 2.0 TDI DSG L1h2 | Дисковые вентилируемые | Дисковые | Гидравлический |
| 2.0 TDI DSG L1h3 | Дисковые вентилируемые | Дисковые | Гидравлический |
| 2.0 TDI DSG L2h2 | Дисковые вентилируемые | Дисковые | Гидравлический |
| 2.0 TDI DSG L2h3 | Дисковые вентилируемые | Дисковые | Гидравлический |
| 2. | Дисковые вентилируемые | Дисковые | Гидравлический |
| 2.0 TDI MT 4Motion L1h2 | Дисковые вентилируемые | Дисковые | Гидравлический |
| 2.0 TDI MT 4Motion L1h3 | Дисковые вентилируемые | Дисковые | Гидравлический |
| 2.0 TDI MT 4Motion L2h2 | Дисковые вентилируемые | Дисковые | Гидравлический |
| 2.0 TDI MT 4Motion L2h3 | Дисковые вентилируемые | Дисковые | Гидравлический |
2. 0 TDI MT 4Motion L2h4 0 TDI MT 4Motion L2h4 | Дисковые вентилируемые | Дисковые | Гидравлический |
| 2.0 TDI MT L1h2 | Дисковые вентилируемые | Дисковые | Гидравлический |
| 2.0 TDI MT L1h3 | Дисковые вентилируемые | Дисковые | Гидравлический |
| 2.0 TDI MT L2h2 | Дисковые вентилируемые | Дисковые | Гидравлический |
| 2.0 TDI MT L2h3 | Дисковые вентилируемые | Дисковые | Гидравлический |
2. 0 TDI MT L2h4 0 TDI MT L2h4 | Дисковые вентилируемые | Дисковые | Гидравлический |
Размер шин
| Модификации | Размер |
|---|---|
| 2.0 BiTDI DSG 4Motion L1h2 | Передние колёса: 235/55 R17, Задние колёса: 235/55 R17 |
| 2.0 BiTDI DSG 4Motion L1h3 | Передние колёса: 235/55 R17, Задние колёса: 235/55 R17 |
| 2.0 BiTDI DSG 4Motion L2h2 | Передние колёса: 235/55 R17, Задние колёса: 235/55 R17 |
| 2.0 BiTDI DSG 4Motion L2h3 | Передние колёса: 235/55 R17, Задние колёса: 235/55 R17 |
| 2.0 BiTDI DSG L1h2 | Передние колёса: 235/55 R17, Задние колёса: 235/55 R17 |
2. 0 BiTDI DSG L1h3 0 BiTDI DSG L1h3 | Передние колёса: 235/55 R17, Задние колёса: 235/55 R17 |
| 2.0 BiTDI DSG L2h2 | Передние колёса: 235/55 R17, Задние колёса: 235/55 R17 |
| 2.0 BiTDI DSG L2h3 | Передние колёса: 235/55 R17, Задние колёса: 235/55 R17 |
| 2.0 BiTDI DSG L2h4 | Передние колёса: 235/55 R17, Задние колёса: 235/55 R17 |
| 2.0 BiTDI MT 4Motion L1h2 | Передние колёса: 235/55 R17, Задние колёса: 235/55 R17 |
| 2.0 BiTDI MT 4Motion L1h3 | Передние колёса: 235/55 R17, Задние колёса: 235/55 R17 |
| 2.0 BiTDI MT 4Motion L2h2 | Передние колёса: 235/55 R17, Задние колёса: 235/55 R17 |
2. 0 BiTDI MT 4Motion L2h3 0 BiTDI MT 4Motion L2h3 | Передние колёса: 235/55 R17, Задние колёса: 235/55 R17 |
| 2.0 BiTDI MT 4Motion L2h4 | Передние колёса: 235/55 R17, Задние колёса: 235/55 R17 |
| 2.0 BiTDI MT L1h2 | Передние колёса: 235/55 R17, Задние колёса: 235/55 R17 |
| 2.0 BiTDI MT L1h3 | Передние колёса: 235/55 R17, Задние колёса: 235/55 R17 |
| 2.0 BiTDI MT L2h2 | Передние колёса: 235/55 R17, Задние колёса: 235/55 R17 |
| 2.0 BiTDI MT L2h3 | Передние колёса: 235/55 R17, Задние колёса: 235/55 R17 |
| 2.0 BiTDI MT L2h4 | Передние колёса: 235/55 R17, Задние колёса: 235/55 R17 |
2. 0 MT L1h2 0 MT L1h2 | Передние колёса: 205/65 R16, Задние колёса: 205/65 R16 |
| 2.0 MT L1h3 | Передние колёса: 205/65 R16, Задние колёса: 205/65 R16 |
| 2.0 MT L2h2 | Передние колёса: 205/65 R16, Задние колёса: 205/65 R16 |
| 2.0 MT L2h3 | Передние колёса: 205/65 R16, Задние колёса: 205/65 R16 |
| 2.0 MT L2h4 | Передние колёса: 205/65 R16, Задние колёса: 205/65 R16 |
| 2.0 TDI DSG L1h2 | Передние колёса: 215/65 R16, Задние колёса: 215/65 R16 |
| 2.0 TDI DSG L1h3 | Передние колёса: 215/65 R16, Задние колёса: 215/65 R16 |
| 2.0 TDI DSG L2h2 | Передние колёса: 215/65 R16, Задние колёса: 215/65 R16 |
2. 0 TDI DSG L2h3 0 TDI DSG L2h3 | Передние колёса: 215/65 R16, Задние колёса: 215/65 R16 |
| 2.0 TDI DSG L2h4 | Передние колёса: 215/65 R16, Задние колёса: 215/65 R16 |
| 2.0 TDI MT 4Motion L1h2 | Передние колёса: 215/65 R16, Задние колёса: 215/65 R16 |
| 2.0 TDI MT 4Motion L1h3 | Передние колёса: 215/65 R16, Задние колёса: 215/65 R16 |
| 2.0 TDI MT 4Motion L2h2 | Передние колёса: 215/65 R16, Задние колёса: 215/65 R16 |
| 2.0 TDI MT 4Motion L2h3 | Передние колёса: 215/65 R16, Задние колёса: 215/65 R16 |
| 2.0 TDI MT 4Motion L2h4 | Передние колёса: 215/65 R16, Задние колёса: 215/65 R16 |
2. 0 TDI MT L1h2 0 TDI MT L1h2 | Передние колёса: 215/65 R16, Задние колёса: 215/65 R16 |
| 2.0 TDI MT L1h3 | Передние колёса: 215/65 R16, Задние колёса: 215/65 R16 |
| 2.0 TDI MT L2h2 | Передние колёса: 215/65 R16, Задние колёса: 215/65 R16 |
| 2.0 TDI MT L2h3 | Передние колёса: 215/65 R16, Задние колёса: 215/65 R16 |
| 2.0 TDI MT L2h4 | Передние колёса: 215/65 R16, Задние колёса: 215/65 R16 |
Размеры
| Модификации | Длина, мм | Ширина, мм | Высота, мм | Колея передняя/задняя, мм | Колесная база, мм | Дорожный просвет (клиренс), мм | Объем багажника, л |
|---|---|---|---|---|---|---|---|
2. 0 BiTDI DSG 4Motion L1h2 0 BiTDI DSG 4Motion L1h2 | 4892 | 1904 | 1990 | -/- | 3000 | 165 | -/- |
| 2.0 BiTDI DSG 4Motion L1h3 | 4892 | 1904 | 2176 | -/- | 3000 | 165 | -/- |
| 2.0 BiTDI DSG 4Motion L2h2 | 5292 | 1904 | 1990 | -/- | 3400 | 165 | -/- |
| 2.0 BiTDI DSG 4Motion L2h3 | 5292 | 1904 | 2176 | -/- | 3400 | 165 | -/- |
2. 0 BiTDI DSG L1h2 0 BiTDI DSG L1h2 | 4892 | 1904 | 1990 | -/- | 3000 | 165 | -/- |
| 2.0 BiTDI DSG L1h3 | 4892 | 1904 | 2176 | -/- | 3000 | 165 | -/- |
| 2.0 BiTDI DSG L2h2 | 5292 | 1904 | 1990 | -/- | 3400 | 165 | -/- |
| 2.0 BiTDI DSG L2h3 | 5292 | 1904 | 2176 | -/- | 3400 | 165 | -/- |
2. 0 BiTDI DSG L2h4 0 BiTDI DSG L2h4 | 5292 | 1904 | 2176 | -/- | 3400 | 165 | -/- |
| 2.0 BiTDI MT 4Motion L1h2 | 4892 | 1904 | 1990 | -/- | 3000 | 165 | -/- |
| 2.0 BiTDI MT 4Motion L1h3 | 4892 | 1904 | 2176 | -/- | 3000 | 165 | -/- |
| 2.0 BiTDI MT 4Motion L2h2 | 5292 | 1904 | 1990 | -/- | 3400 | 165 | -/- |
2. 0 BiTDI MT 4Motion L2h3 0 BiTDI MT 4Motion L2h3 | 5292 | 1904 | 2176 | -/- | 3400 | 165 | -/- |
| 2.0 BiTDI MT 4Motion L2h4 | 5292 | 1904 | 2176 | -/- | 3400 | 165 | -/- |
| 2.0 BiTDI MT L1h2 | 4892 | 1904 | 1990 | -/- | 3000 | 165 | -/- |
| 2.0 BiTDI MT L1h3 | 4892 | 1904 | 2176 | -/- | 3000 | 165 | -/- |
2. 0 BiTDI MT L2h2 0 BiTDI MT L2h2 | 5292 | 1904 | 1990 | -/- | 3400 | 165 | -/- |
| 2.0 BiTDI MT L2h3 | 5292 | 1904 | 2176 | -/- | 3400 | 165 | -/- |
| 2.0 BiTDI MT L2h4 | 5292 | 1904 | 2176 | -/- | 3400 | 165 | -/- |
| 2.0 MT L1h2 | 4892 | 1904 | 1990 | -/- | 3000 | 165 | -/- |
2. 0 MT L1h3 0 MT L1h3 | 4892 | 1904 | 2176 | -/- | 3000 | 165 | -/- |
| 2.0 MT L2h2 | 5292 | 1904 | 1990 | -/- | 3400 | 165 | -/- |
| 2.0 MT L2h3 | 5292 | 1904 | 2176 | -/- | 3400 | 165 | -/- |
| 2.0 MT L2h4 | 5292 | 1904 | 2176 | -/- | 3400 | 165 | -/- |
2. 0 TDI DSG L1h2 0 TDI DSG L1h2 | 4892 | 1904 | 1990 | -/- | 3000 | 165 | -/- |
| 2.0 TDI DSG L1h3 | 4892 | 1904 | 2176 | -/- | 3000 | 165 | -/- |
| 2.0 TDI DSG L2h2 | 5292 | 1904 | 1990 | -/- | 3400 | 165 | -/- |
| 2.0 TDI DSG L2h3 | 5292 | 1904 | 2176 | -/- | 3400 | 165 | -/- |
2. 0 TDI DSG L2h4 0 TDI DSG L2h4 | 5292 | 1904 | 2176 | -/- | 3400 | 165 | -/- |
| 2.0 TDI MT 4Motion L1h2 | 4892 | 1904 | 1990 | -/- | 3000 | 165 | -/- |
| 2.0 TDI MT 4Motion L1h3 | 4892 | 1904 | 2176 | -/- | 3000 | 165 | -/- |
| 2.0 TDI MT 4Motion L2h2 | 5292 | 1904 | 1990 | -/- | 3400 | 165 | -/- |
2. 0 TDI MT 4Motion L2h3 0 TDI MT 4Motion L2h3 | 5292 | 1904 | 2176 | -/- | 3400 | 165 | -/- |
| 2.0 TDI MT 4Motion L2h4 | 5292 | 1904 | 2176 | -/- | 3400 | 165 | -/- |
| 2.0 TDI MT L1h2 | 4892 | 1904 | 1990 | -/- | 3000 | 165 | -/- |
| 2.0 TDI MT L1h3 | 4892 | 1904 | 2176 | -/- | 3000 | 165 | -/- |
| 2. | 5292 | 1904 | 1990 | -/- | 3400 | 165 | -/- |
| 2.0 TDI MT L2h3 | 5292 | 1904 | 2176 | -/- | 3400 | 165 | -/- |
| 2.0 TDI MT L2h4 | 5292 | 1904 | 2176 | -/- | 3400 | 165 | -/- |
Вес автомобиля
| Модификации | Снаряженная масса, кг | Максимальная масса, кг | Грузоподъёмность, кг |
|---|---|---|---|
2. 0 BiTDI DSG 4Motion L1h2 0 BiTDI DSG 4Motion L1h2 | 1978 | 2800 | 822 |
| 2.0 BiTDI DSG 4Motion L1h3 | 1978 | 2800 | 822 |
| 2.0 BiTDI DSG 4Motion L2h2 | 2029 | 2800 | 771 |
| 2.0 BiTDI DSG 4Motion L2h3 | 2029 | 2800 | 771 |
| 2.0 BiTDI DSG L1h2 | 1856 | 2800 | 944 |
| 2.0 BiTDI DSG L1h3 | 1856 | 2800 | 944 |
| 2. | 1907 | 2800 | 893 |
| 2.0 BiTDI DSG L2h3 | 1907 | 2800 | 893 |
| 2.0 BiTDI DSG L2h4 | 1907 | 2800 | 893 |
| 2.0 BiTDI MT 4Motion L1h2 | 1844 | 2800 | 956 |
| 2.0 BiTDI MT 4Motion L1h3 | 1844 | 2800 | 956 |
| 2.0 BiTDI MT 4Motion L2h2 | — | — | — |
2. 0 BiTDI MT 4Motion L2h3 0 BiTDI MT 4Motion L2h3 | — | 2800 | 2800 |
| 2.0 BiTDI MT 4Motion L2h4 | — | 2800 | 2800 |
| 2.0 BiTDI MT L1h2 | 1844 | 2800 | 956 |
| 2.0 BiTDI MT L1h3 | 1844 | 2800 | 956 |
| 2.0 BiTDI MT L2h2 | 1895 | 2800 | 905 |
| 2.0 BiTDI MT L2h3 | 1895 | 2800 | 905 |
2. 0 BiTDI MT L2h4 0 BiTDI MT L2h4 | 1895 | 2800 | 905 |
| 2.0 MT L1h2 | 1729 | 2600 | 871 |
| 2.0 MT L1h3 | 1729 | 2850 | 1121 |
| 2.0 MT L2h2 | 1780 | 2850 | 1070 |
| 2.0 MT L2h3 | 1780 | 2850 | 1070 |
| 2.0 MT L2h4 | 1780 | 2850 | 1070 |
| 2.0 TDI DSG L1h2 | 1809 | 2800 | 991 |
2. 0 TDI DSG L1h3 0 TDI DSG L1h3 | 1809 | 2800 | 991 |
| 2.0 TDI DSG L2h2 | 1860 | 2800 | 940 |
| 2.0 TDI DSG L2h3 | 1860 | 2800 | 940 |
| 2.0 TDI DSG L2h4 | 1860 | 2800 | 940 |
| 2.0 TDI MT 4Motion L1h2 | 1917 | 2800 | 883 |
| 2.0 TDI MT 4Motion L1h3 | 1917 | 2800 | 883 |
2. 0 TDI MT 4Motion L2h2 0 TDI MT 4Motion L2h2 | 1968 | 2800 | 832 |
| 2.0 TDI MT 4Motion L2h3 | 1968 | 2800 | 832 |
| 2.0 TDI MT 4Motion L2h4 | 1968 | 2800 | 832 |
| 2.0 TDI MT L1h2 | 1797 | 2800 | 1003 |
| 2.0 TDI MT L1h3 | 1797 | 2800 | 1003 |
| 2.0 TDI MT L2h2 | 1797 | 2800 | 1003 |
2. 0 TDI MT L2h3 0 TDI MT L2h3 | 1797 | 2800 | 1003 |
| 2.0 TDI MT L2h4 | 1797 | 2800 | 1003 |
Количество сидений и дверей
| Модификации | Количество сидений | Количество дверей |
|---|---|---|
| 2.0 BiTDI DSG 4Motion L1h2 | 2 | 4 |
| 2.0 BiTDI DSG 4Motion L1h3 | 2 | 4 |
| 2.0 BiTDI DSG 4Motion L2h2 | 2 | 4 |
| 2.0 BiTDI DSG 4Motion L2h3 | 2 | 4 |
2. 0 BiTDI DSG L1h2 0 BiTDI DSG L1h2 | 2 | 4 |
| 2.0 BiTDI DSG L1h3 | 2 | 4 |
| 2.0 BiTDI DSG L2h2 | 2 | 4 |
| 2.0 BiTDI DSG L2h3 | 2 | 4 |
| 2.0 BiTDI DSG L2h4 | 2 | 4 |
| 2.0 BiTDI MT 4Motion L1h2 | 2 | 4 |
| 2.0 BiTDI MT 4Motion L1h3 | 2 | 4 |
| 2.0 BiTDI MT 4Motion L2h2 | 2 | 4 |
| 2. | 2 | 4 |
| 2.0 BiTDI MT 4Motion L2h4 | 2 | 4 |
| 2.0 BiTDI MT L1h2 | 2 | 4 |
| 2.0 BiTDI MT L1h3 | 2 | 4 |
| 2.0 BiTDI MT L2h2 | 2 | 4 |
| 2.0 BiTDI MT L2h3 | 2 | 4 |
| 2.0 BiTDI MT L2h4 | 2 | 4 |
| 2.0 MT L1h2 | 2 | 4 |
2..jpg) 0 MT L1h3 0 MT L1h3 | 2 | 4 |
| 2.0 MT L2h2 | 2 | 4 |
| 2.0 MT L2h3 | 2 | 4 |
| 2.0 MT L2h4 | 2 | 4 |
| 2.0 TDI DSG L1h2 | 2 | 4 |
| 2.0 TDI DSG L1h3 | 2 | 4 |
| 2.0 TDI DSG L2h2 | 2 | 4 |
| 2.0 TDI DSG L2h3 | 2 | 4 |
| 2.0 TDI DSG L2h4 | 2 | 4 |
2. 0 TDI MT 4Motion L1h2 0 TDI MT 4Motion L1h2 | 2 | 4 |
| 2.0 TDI MT 4Motion L1h3 | 2 | 4 |
| 2.0 TDI MT 4Motion L2h2 | 2 | 4 |
| 2.0 TDI MT 4Motion L2h3 | 2 | 4 |
| 2.0 TDI MT 4Motion L2h4 | 2 | 4 |
| 2.0 TDI MT L1h2 | 2 | 4 |
| 2.0 TDI MT L1h3 | 2 | 4 |
| 2.0 TDI MT L2h2 | 2 | 4 |
2. 0 TDI MT L2h3 0 TDI MT L2h3 | 2 | 4 |
| 2.0 TDI MT L2h4 | 2 | 4 |
Объём топливного бака
| Модификации | Объём бака, л | Тип топлива |
|---|---|---|
| 2.0 BiTDI DSG 4Motion L1h2 | 80 | ДТ |
| 2.0 BiTDI DSG 4Motion L1h3 | 80 | ДТ |
| 2.0 BiTDI DSG 4Motion L2h2 | 80 | ДТ |
| 2.0 BiTDI DSG 4Motion L2h3 | 80 | ДТ |
| 2.0 BiTDI DSG L1h2 | 80 | ДТ |
2. 0 BiTDI DSG L1h3 0 BiTDI DSG L1h3 | 80 | ДТ |
| 2.0 BiTDI DSG L2h2 | 80 | ДТ |
| 2.0 BiTDI DSG L2h3 | 80 | ДТ |
| 2.0 BiTDI DSG L2h4 | 80 | ДТ |
| 2.0 BiTDI MT 4Motion L1h2 | 80 | ДТ |
| 2.0 BiTDI MT 4Motion L1h3 | 80 | ДТ |
| 2.0 BiTDI MT 4Motion L2h2 | 80 | ДТ |
| 2.0 BiTDI MT 4Motion L2h3 | 80 | ДТ |
| 2.0 BiTDI MT 4Motion L2h4 | 80 | ДТ |
2. 0 BiTDI MT L1h2 0 BiTDI MT L1h2 | 80 | ДТ |
| 2.0 BiTDI MT L1h3 | 80 | ДТ |
| 2.0 BiTDI MT L2h2 | 80 | ДТ |
| 2.0 BiTDI MT L2h3 | 80 | ДТ |
| 2.0 BiTDI MT L2h4 | 80 | ДТ |
| 2.0 MT L1h2 | 80 | АИ-95 |
| 2.0 MT L1h3 | 80 | АИ-95 |
| 2.0 MT L2h2 | 80 | АИ-95 |
| 2.0 MT L2h3 | 80 | АИ-95 |
2. 0 MT L2h4 0 MT L2h4 | 80 | АИ-95 |
| 2.0 TDI DSG L1h2 | 80 | ДТ |
| 2.0 TDI DSG L1h3 | 80 | ДТ |
| 2.0 TDI DSG L2h2 | 80 | ДТ |
| 2.0 TDI DSG L2h3 | 80 | ДТ |
| 2.0 TDI DSG L2h4 | 80 | ДТ |
| 2.0 TDI MT 4Motion L1h2 | 80 | ДТ |
| 2.0 TDI MT 4Motion L1h3 | 80 | ДТ |
| 2.0 TDI MT 4Motion L2h2 | 80 | ДТ |
2. 0 TDI MT 4Motion L2h3 0 TDI MT 4Motion L2h3 | 80 | ДТ |
| 2.0 TDI MT 4Motion L2h4 | 80 | ДТ |
| 2.0 TDI MT L1h2 | 80 | ДТ |
| 2.0 TDI MT L1h3 | 80 | ДТ |
| 2.0 TDI MT L2h2 | 80 | ДТ |
| 2.0 TDI MT L2h3 | 80 | ДТ |
| 2.0 TDI MT L2h4 | 80 | ДТ |
Динамика
| Модификации | Максимальная скорость, км/ч | Время разгона до 100 км/ч, с | Cd (Коэффициент лобового сопротивления) |
|---|---|---|---|
| 2. | 188 | 10.6 | — |
| 2.0 BiTDI DSG 4Motion L1h3 | 182 | 10.7 | — |
| 2.0 BiTDI DSG 4Motion L2h2 | 188 | 10.6 | — |
| 2.0 BiTDI DSG 4Motion L2h3 | 182 | 10.7 | — |
| 2.0 BiTDI DSG L1h2 | 191 | 10 | — |
| 2.0 BiTDI DSG L1h3 | 184 | 10.1 | — |
| 2.0 BiTDI DSG L2h2 | 191 | 10 | — |
2. 0 BiTDI DSG L2h3 0 BiTDI DSG L2h3 | 184 | 10.1 | — |
| 2.0 BiTDI DSG L2h4 | 176 | 10.4 | — |
| 2.0 BiTDI MT 4Motion L1h2 | — | — | — |
| 2.0 BiTDI MT 4Motion L1h3 | — | — | — |
| 2.0 BiTDI MT 4Motion L2h2 | — | — | — |
| 2.0 BiTDI MT 4Motion L2h3 | — | — | — |
| 2.0 BiTDI MT 4Motion L2h4 | — | — | — |
2. 0 BiTDI MT L1h2 0 BiTDI MT L1h2 | 191 | 9.9 | — |
| 2.0 BiTDI MT L1h3 | 184 | 10 | — |
| 2.0 BiTDI MT L2h2 | 191 | 9.9 | — |
| 2.0 BiTDI MT L2h3 | 184 | 10 | — |
| 2.0 BiTDI MT L2h4 | 176 | 10.3 | — |
| 2.0 MT L1h2 | 163 | 15 | — |
| 2.0 MT L1h3 | 158 | 15.1 | — |
2. 0 MT L2h2 0 MT L2h2 | 163 | 15 | — |
| 2.0 MT L2h3 | 158 | 15.1 | — |
| 2.0 MT L2h4 | 152 | 15.6 | — |
| 2.0 TDI DSG L1h2 | 172 | 12.5 | — |
| 2.0 TDI DSG L1h3 | 172 | 12.5 | — |
| 2.0 TDI DSG L2h2 | 172 | 12.5 | — |
| 2.0 TDI DSG L2h3 | 172 | 12.5 | — |
| 2. | 160 | 13 | — |
| 2.0 TDI MT 4Motion L1h2 | 170 | 13.5 | — |
| 2.0 TDI MT 4Motion L1h3 | 164 | 13.6 | — |
| 2.0 TDI MT 4Motion L2h2 | 170 | 13.5 | — |
| 2.0 TDI MT 4Motion L2h3 | 164 | 13.6 | — |
| 2.0 TDI MT 4Motion L2h4 | 158 | 14 | — |
| 2.0 TDI MT L1h2 | 173 | 12. 2 2 | — |
| 2.0 TDI MT L1h3 | 167 | 12.3 | — |
| 2.0 TDI MT L2h2 | 173 | 12.2 | — |
| 2.0 TDI MT L2h3 | 167 | 12.3 | — |
| 2.0 TDI MT L2h4 | 161 | 12.7 | — |
Расход топлива
| Модификации | В городе, л/100 км | По трассе, л/100 км | Средний расход, л/100 км | Выброс СО2, г/км | Тип топлива |
|---|---|---|---|---|---|
2. 0 BiTDI DSG 4Motion L1h2 0 BiTDI DSG 4Motion L1h2 | 10.8 | 7.2 | 8.5 | — | Дизель |
| 2.0 BiTDI DSG 4Motion L1h3 | 10.8 | 7.2 | 8.5 | — | Дизель |
| 2.0 BiTDI DSG 4Motion L2h2 | 10.8 | 7.2 | 8.5 | — | Дизель |
| 2.0 BiTDI DSG 4Motion L2h3 | 10.8 | 7.2 | 8.5 | — | Дизель |
| 2.0 BiTDI DSG L1h2 | 9.9 | 6. 6 6 | 7.8 | — | Дизель |
| 2.0 BiTDI DSG L1h3 | 9.9 | 6.6 | 7.8 | — | Дизель |
| 2.0 BiTDI DSG L2h2 | 9.9 | 6.6 | 7.8 | — | Дизель |
| 2.0 BiTDI DSG L2h3 | 9.9 | 6.6 | 7.8 | — | Дизель |
| 2.0 BiTDI DSG L2h4 | 9.9 | 7 | 8.1 | — | Дизель |
2. 0 BiTDI MT L1h2 0 BiTDI MT L1h2 | 9.5 | 6.4 | 7.5 | — | Дизель |
| 2.0 BiTDI MT L1h3 | 9.5 | 6.4 | 7.5 | — | Дизель |
| 2.0 BiTDI MT L2h2 | 9.5 | 6.4 | 7.5 | — | Дизель |
| 2.0 BiTDI MT L2h3 | 9.5 | 6.4 | 7.5 | — | Дизель |
| 2.0 BiTDI MT L2h4 | 9.5 | 6.8 | 7. | — | Дизель |
| 2.0 MT L1h2 | 13.5 | 8.5 | 10.3 | — | Бензин |
| 2.0 MT L1h3 | 13.5 | 8.5 | 10.3 | — | Бензин |
| 2.0 MT L2h2 | 13.5 | 8.5 | 10.3 | — | Бензин |
| 2.0 MT L2h3 | 13.5 | 8.5 | 10.3 | — | Бензин |
| 2.0 MT L2h4 | 13. 7 | 8.8 | 10.6 | — | Бензин |
| 2.0 TDI DSG L1h2 | 10.1 | 6.6 | 7.9 | — | Дизель |
| 2.0 TDI DSG L1h3 | 10.1 | 6.6 | 7.9 | — | Дизель |
| 2.0 TDI DSG L2h2 | 10.1 | 6.6 | 7.9 | — | Дизель |
| 2.0 TDI DSG L2h3 | 10.1 | 6.6 | 7.9 | — | Дизель |
| 2. 0 TDI DSG L2h4 | 10.1 | 7.1 | 8.2 | — | Дизель |
| 2.0 TDI MT 4Motion L1h2 | 10.1 | 6.8 | 8 | — | Дизель |
| 2.0 TDI MT 4Motion L1h3 | 10.1 | 6.8 | 8 | — | Дизель |
| 2.0 TDI MT 4Motion L2h2 | 10.1 | 6.8 | 8 | — | Дизель |
| 2.0 TDI MT 4Motion L2h3 | 10.1 | 6.8 | 8 | — | Дизель |
| 2. 0 TDI MT 4Motion L2h4 | 10.1 | 7.2 | 8.3 | — | Дизель |
| 2.0 TDI MT L1h2 | 9.5 | 6.2 | 7.4 | — | Дизель |
| 2.0 TDI MT L1h3 | 9.5 | 6.2 | 7.4 | — | Дизель |
| 2.0 TDI MT L2h2 | 9.5 | 6.2 | 7.4 | — | Дизель |
| 2.0 TDI MT L2h3 | 9.5 | 6.2 | 7.4 | — | Дизель |
| 2. 0 TDI MT L2h4 | 9.9 | 6.6 | 7.8 | — | Дизель |
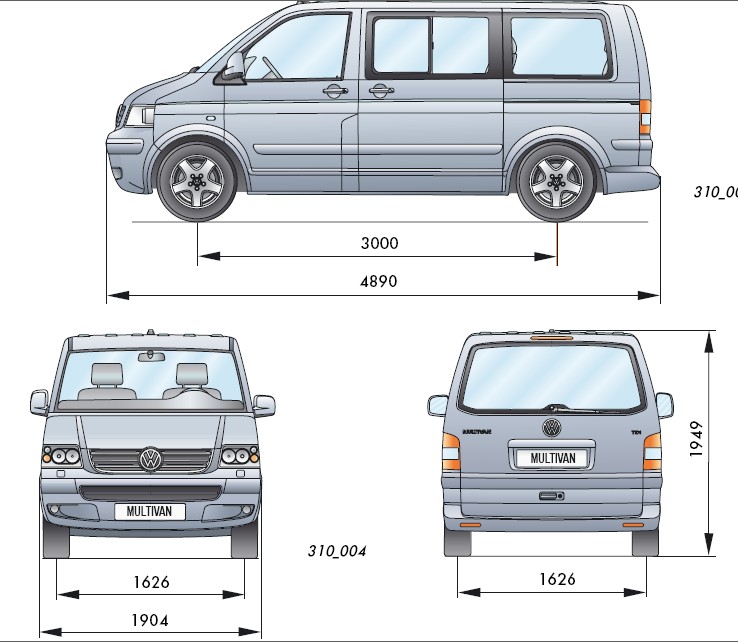
2003 Volkswagen Multivan (T5) 2.5 TDI (174 лс)
2003 Volkswagen Multivan (T5) 2.5 TDI (174 лс) | Технические характеристики, расход топлива , ГабаритыГлавная >> Авто каталог >> Volkswagen >> Multivan >> 2003 Multivan (T5) >> 2.5 TDI (174 лс)
Авто каталог Логин Регистрация Добавить автомобиль Car Specs API
Volkswagen Volkswagen Multivan 2003 Multivan (T5) 3.2 V6 (235 лс) Tiptronic 2.5 TDI (174 лс)
| Какой тип кузова, Volkswagen Multivan (T5)? | Минивэн, 4 дверей, 7 мест |
|---|---|
| Какой расход топлива, Volkswagen Multivan (T5) 2. 5 TDI (174 Hp)? | 8.3 л/100 км 28.34 US mpg 34.03 UK mpg 12.05 км/л |
| Насколько ЭКОЛОГИЧЕН автомобиль, Volkswagen Multivan 2.5 TDI (174 Hp)? | 221 г/км CO2 Euro 3 |
| Насколько быстра машина, 2003 Multivan (T5) 2.5 TDI (174 Hp)? | 183 км/ч | 113.71 mph 0-100 км/ч: 12.2 сек 0-60 mph: 11.6 сек |
| Какая мощность двигателя, Volkswagen Multivan Минивэн 2003 2.5 TDI (174 Hp)? | 174 лс, 340 Нм 250.77 lb.-ft. |
| Какой объем двигателя, Volkswagen Multivan Минивэн 2003 2.5 TDI (174 Hp)? | 2.5 л 2461 см3 150.18 cu. in. |
| Сколько цилиндров имеет мотор, 2003 Volkswagen 2.5 TDI (174 Hp)? | 5, Рядный |
| Какой привод у автомобиля, Volkswagen Multivan (T5) Минивэн 2003 2.5 TDI (174 Hp)? | Передний привод. Двигатель внутреннего сгорания. ДВС приводит в движение передние колеса автомобиля. |
| Какая длина автомобиля, 2003 Volkswagen Multivan Минивэн? | 4890 мм 192. 52 in. |
| Насколько широка машина, 2003 Volkswagen Multivan Минивэн? | 1904 мм 74.96 in. |
| Сколько весит автомобиль, 2003 Volkswagen Multivan (T5) 2.5 TDI (174 Hp)? | 2349 кг 5178.66 lbs. |
| Какая максимально допустимая масса, 2003 Volkswagen Multivan (T5) 2.5 TDI (174 Hp)? | 3000 кг 6613.87 lbs. |
| Сколько передач имеет коробка, Какой тип коробки передач, 2003 Volkswagen Multivan (T5) 2.5 TDI (174 Hp)? | 6, Механическая коробка передач |
- Volkswagen
- Volkswagen Multivan
- 2003 Multivan (T5)
- 3.2 V6 (235 лс) Tiptronic
- 2.5 TDI (174 лс)
- 2003 Multivan (T5)
- Volkswagen Multivan
AcuraAlfa RomeoAlpinaAston MartinAudiBentleyBMWBugattiCadillacChevroletChryslerCitroenCupraDaciaDaewooDaihatsuDodgeDSFerrariFiatFordGMCGreat WallHavalHondaHummerHyundaiInfinitiJaguarJeepKiaKoenigseggLadaLamborghiniLanciaLand RoverLexusLotusMaseratiMazdaMcLarenMercedes-BenzMGMiniMitsubishiNIONissanOpelPaganiPeugeotPorscheRAMRenaultRolls-RoyceRoverSaabSeatSkodaSmartSubaruSuzukiTeslaToyotaVauxhallVolkswagenVolvo
Все бренды
Сравнение
| Базовая информация | |
|---|---|
| Марка | Volkswagen |
| Модель | Multivan |
| Поколения | Multivan (T5) |
| Модификация (двигатель) | 2.5 TDI (174 лс) |
| Начало выпуска | 2003 г |
| Оконч. выпуска | 2006 г |
| Архитектура силового агрегата | Двигатель внутреннего сгорания |
| Тип кузова | Минивэн |
| Количество мест | 7 |
| Количество дверей | 4 |
| Эксплуатационные характеристики | |
| Расход топлива в городе | 11 л/100 км
21.38 US mpg 25.68 UK mpg 9.09 км/л |
| Расход топлива на шоссе | 6.8 л/100 км
34.59 US mpg 41.54 UK mpg 14.71 км/л |
| Расход топлива Смешанный цикл | 8. 3 л/100 км
28.34 US mpg 34.03 UK mpg 12.05 км/л |
| Выбросы CO2 | 221 г/км |
| Топливо | Дизельное топливо |
| Время разгона 0 — 100 км/ч | 12.2 сек |
| Время разгона 0 — 62 mph | 12.2 сек |
| Время разгона 0 — 60 mph (Рассчитано Auto-Data.net) | 11.6 сек |
| Максимальная скорость | 183 км/ч 113.71 mph |
| Экологический стандарт | Euro 3 |
| Соотношение мощность/вес | 13.5 кг/лс, 74.1 лс/тонна |
| Соотношение Крутящий момент/вес | 6.9 кг/Нм, 144.7 Нм/тонна |
| Двигатель | |
| Мощность | 174 лс @ 3500 об./мин. |
| Мощность на литр рабочего объема | 70.7 лс/л |
| Крутящий момент | 340 Нм @ 2000 об./мин.
250.77 lb.-ft. @ 2000 об. /мин. |
| Расположение двигателя | переднее, продольное |
| Модель/Код двигателя | AXE |
| Объем двигателя | 2461 см3 150.18 cu. in. |
| Количество цилиндров | 5 |
| Расположение цилиндров | Рядный |
| Диаметр цилиндра | 81 мм 3.19 in. |
| Ход поршня | 95.5 мм 3.76 in. |
| Степень сжатия | 18.5 |
| Количество клапанов на цилиндр | 2 |
| Система питания | Помпа-дюзе |
| Тип наддува | Турбонаддув, Интеркулер |
| Количество масла в двигателе | 7.4 л 7.82 US qt | 6.51 UK qt |
| Спецификация моторного масла | Войдите, чтобы увидеть. |
| охлаждающая жидкость | 7.1 л
7.5 US qt | 6. 25 UK qt |
| Объем и вес | |
| Снаряженная масса автомобиля | 2349 кг 5178.66 lbs. |
| Допустимая полная масса | 3000 кг 6613.87 lbs. |
| Максимальная грузоподъемность | 651 кг 1435.21 lbs. |
| Объем топливного бака | 80 л 21.13 US gal | 17.6 UK gal |
| Максимальная нагрузка на крышу | 100 кг 220.46 lbs. |
| Допустимая масса буксируемого прицепа при уклоне 12% | 2500 кг 5511.56 lbs. |
| Допустимая масса буксируемого прицепа без тормозной системы | 750 кг 1653.47 lbs. |
| Габариты | |
| Длина | 4890 мм 192.52 in. |
| Ширина | 1904 мм
74. 96 in. |
| Высота | 1970 мм 77.56 in. |
| Колесная база | 3400 мм 133.86 in. |
| Колея передняя | 1628 мм 64.09 in. |
| Колея задняя | 1628 мм 64.09 in. |
| Диаметр разворота | 11.9 м 39.04 ft. |
| Трансмиссия, тормоза и подвеска | |
| Архитектура привода | ДВС приводит в движение передние колеса автомобиля. |
| Привод | Передний привод |
| Количество передач (Механическая коробка передач) | 6 |
| Размер шин | 215/65 R16 |
| Размер дисков | 16 |
Какие доступны лампы T5? | Флуоресцентные системы T5 | Освещение ответы
Какие лампы T5 доступны?Лампы T5 доступны для стандартной и высокой мощности. В Таблице 1-3 показаны характеристики четырехфутовых стандартных ламп T8 и T12. Стандартные лампы T5 и T5 HO имеют расчетный средний срок службы 20 000 часов, такой же, как у большинства стандартных ламп T8 и T12. Новые лампы T8 и T12 с увеличенным сроком службы имеют срок службы на 4 000–10 000 часов больше, чем лампы T5. (Таблица 1-3 не включает эти лампы.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Идентификация и характеристика T5-подобных бактериофагов, представляющих две новые подгруппы из пищевых продуктов
- Список журналов
- Фронт микробиол
- PMC5816814
Передняя микробиол. 2018; 9: 202.
Опубликовано в Интернете 13 февраля 2018 г. doi: 10.3389/fmicb.2018.00202
, 1 , 2 , 3 , 4 , 2, * и 1, *
Авторская информация Примечания Copyright и лицензионная информация о отказе
- Дополнительные материалы. увеличилась заболеваемость возбудителями пищевых инфекций, особенно представителей семейства Enterobacteriaceae, среди которых патогенные штаммы Escherichia coli, Shigella и Salmonella . Здесь мы сообщаем о выделении и характеристике 12 новых T5-подобных бактериофагов из конфискованных образцов пищевых продуктов. Все бактериофаги эффективно лизируются штаммов E. coli K-12 и были способны инфицировать патогенные штаммы E. coli , представляющие энтерогеморрагический (EHEC), энтеропатогенный (EPEC), энтеротоксигенный (ETEC) и энтероинвазивный (EIEC) патотипы, Shigella dysenteriae, S. sonnei , а также штаммы E. coli с множественной лекарственной устойчивостью (MDR) и несколько штаммов, представляющих разные серовары Salmonella enterica . Все бактериофаги имели морфологию Siphoviridae . Секвенирование всего генома новых T5-подобных бактериофагов показало, что они представляют две отдельные группы, при этом группировка на основе генома коррелирует с различными спектрами хозяев. Поскольку эти бактериофаги имеют пищевое происхождение, их стабильность и отсутствие каких-либо генов вирулентности, а также их широкий и взаимодополняющий спектр хозяев делают эти новые Т5-подобные бактериофаги ценными кандидатами для использования в качестве агентов биологической борьбы с патогенными энтеробактериями пищевого происхождения.
Ключевые слова: бактериофаги, T5-подобные фаги, фаговая геномика, Enterobacteriaceae, ESBL E. coli , безопасность пищевых продуктов, биоконтроль (EHEC) и штаммов Shigella , способных вызывать опасные для жизни заболевания даже при очень небольшой инфекционной дозе (обзор в Croxen et al., 2013; Anderson et al., 2016). Многие из этих инфекций лечат с помощью антибиотикотерапии, но этому угрожает рост числа штаммов с множественной лекарственной устойчивостью (МЛУ) (см. обзор Szmolka and Nagy, 2013). Разработка новых антибиотиков значительно замедлилась в последние десятилетия, поскольку становится все труднее найти агенты с новыми механизмами действия или мишенями, а также из-за ограниченной экономической отдачи (Hunter, 2012). Совсем недавно внимание было обращено на разработку антибиотиков, «устойчивых к эволюции» (Bell and MacLean, 2017), и в качестве альтернативного ответа на этот вызов также возродился интерес к изучению бактериофагов, способных лизировать патогенные бактерии.
В качестве агентов биологической борьбы фаги могут применяться либо для лечения продолжающихся инфекций (фаговая терапия), как в случае штаммов Staphylococcus aureus , либо для патогенов пищевого происхождения, обработка пищевых продуктов в качестве профилактической меры (рассмотрено Johnson и др., 2008 г. и Хагенс и Лесснер, 2010 г.). Уже есть несколько примеров коммерчески доступных фаговых коктейлей, которые можно использовать против различных патогенов пищевого происхождения. Существует фаговый коктейль, направленный против E. coli O157 для нанесения на пищевые поверхности (Carter et al., 2012), а также коктейли против Shigella (Soffer et al., 2017), Listeria monocytogenes и Salmonella enterica (отзыв Абедон, 2017). Также проводились клинические испытания фаговых коктейлей против диареегенной E. coli на людях (Sarker et al., 2012, 2016), но они не дали результатов в отношении эффективности использования фаговых коктейлей при таких инфекциях.
Бактериофаг T5 был одним из первых энтеробактериальных фагов, подробно охарактеризованных в 1950-х годах, и с тех пор было сообщено о нескольких подобных бактериофагах. Их наиболее уникальной особенностью является двухэтапный перенос ДНК во время инфекции (обзор в Davison, 2015). Позже, отчасти из-за возобновившегося интереса к использованию бактериофагов в качестве антибактериальных агентов, был полностью секвенирован геном бактериофага Т5 (Wang et al., 2005), а также были охарактеризованы новые члены этой группы бактериофагов (Kim and Ryu, 2011; Golomidova et al. др., 2015). Все бактериофаги T5, по-видимому, имеют общую геномную структуру, в которой можно выделить преранние, ранние и поздние области в зависимости от времени транскрипции в течение инфекционного цикла (обзор Davison, 2015). В недавних исследованиях несколько T5-подобных бактериофагов были предложены в качестве потенциальных кандидатов для использования против различных патогенов пищевого происхождения, в том числе Salmonella enterica серовара Typhimurium (Kim and Ryu, 2011; Piya et al.
, 2015), а также против штаммов E. coli O157:H7 (Niu et al., 2012; Hong et al., 2014), с многообещающие результаты in vivo , полученные на живых овцах с помощью фага CEV2 (Raya et al., 2011).В предыдущем исследовании мы оценили риск, связанный с пищевыми патогенами, присутствующими в нелегально ввезенных в Европу пищевых продуктах, с особым акцентом на STEC (Nagy et al., 2015). Мы предположили, что бактериофаги, способные лизировать патогенные бактерии пищевого происхождения, также должны быть способны выживать и размножаться в той же пище. Таким образом, те же самые образцы пищевых продуктов могут стать мощным источником кандидатов на бактериофаги, которые можно применять в качестве средства биологической борьбы с пищевыми бактериями. Как указал Abedon (2017), исследования, направленные на фаговую терапию, как правило, сосредоточены на успехах в терапевтическом применении и пренебрегают подробной характеристикой фагов. Исследования потенциального терапевтического применения фагов против энтеробактериальных инфекций (Sarker et al.
, 2016) показывают, что профилактический подход, то есть предварительное уничтожение патогенов из пищи с помощью фагов перед употреблением, может быть более успешным применением.В свете этих соображений мы стремились изолировать бактериофаги из образцов пищевых продуктов, которые способны лизировать кишечные патогены пищевого происхождения, чтобы подробно охарактеризовать спектр их хозяев с другими фенотипическими признаками и охарактеризовать их геномы. Эта всесторонняя характеристика послужит для определения того, можно ли рассматривать выделенные фаги в качестве агентов биологической борьбы с патогенными энтеробактериями, переносимыми через пищу.
Используя штаммы E. coli K-12 для размножения, мы выделили 12 новых T5-подобных литических бактериофагов из отдельных образцов пищевых продуктов и приступили к их подробной характеристике. Анализ последовательности всего генома показал, что эти бактериофаги не содержат нежелательных генов, связанных с вирулентностью.
Они представляют собой два разных новых генотипа среди Т5-подобных бактериофагов и доказали свою способность лизировать широкий спектр патогенных штаммов E. coli, Shigella и Salmonella .Выделение бактериофагов
Бактериофаги были выделены из образцов пищевых продуктов, конфискованных на венгерской границе таможенниками. Образцы прошли первые этапы метода ISO 16654:2001 для выделения E. coli O157. Вкратце, 5 г образцов пищевых продуктов гомогенизировали в 10 объемах триптического соевого бульона (ТСБ) с добавлением солей желчных кислот и инкубировали в течение 24 часов при 42°С. Безбактериальные образцы этих прекультур наносили или наносили пятнами на слоистую чашку с 0,7% мягким агаром, содержащую E. coli Производное К-12 С600 в качестве размножающегося штамма, как описано Strauch et al. (2001). После инкубации в течение ночи при 37°C отдельные бляшки собирали и очищали повторным размножением на E. coli C600, а также на E.
coli MG1655 не менее трех раз до достижения высокого титра (≥10 11 PFU). /мл) были получены запасы бактериофагов. Список выделенных бактериофагов и их происхождение приведены в табл.Таблица 1
Происхождение и подгруппы новых T5-подобных бактериофагов.
Фаг № . Foodstuff of origin Country of origin Subgroup chee24 cattle cheese Bulgaria chee24 pork27 row pork meat Serbia сыр24 свинина29 рядовая свинина Сербия сыр24 saus47N pork sausage Serbia chee24 saus111K pork sausage Ukraine chee24 poul124 poultry meat Ukraine chee24 chee130_1 сыр КРС Украина сыр130_1 колбаса132 колбаса свиная Украина chee130_1 poul149 poultry meat Ukraine chee130_1 chee158 cattle cheese Ukraine chee130_1 cott162 cattle cottage cheese Ukraine chee130_1 saus176 N Колбаса свиная Венгрия chee130_1 Открыть в отдельном окне
Проверка специфичности к хозяину и эффективности посева
Специфичность выделенных бактериофагов к хозяину исследовали с помощью точечных анализов штаммов, перечисленных в таблице, как описано выше.
В дополнение к эталонным штаммам и штаммам из нашей коллекции штаммов были исследованы штаммы E. coli , продуцирующие бета-лактамазы расширенного спектра действия (БЛРС), с множественной лекарственной устойчивостью (МЛУ), выделенные из клинических образцов человека. Штаммы MDR были выделены в Университете Дебрецена и 9Штаммы 0457 Salmonella были любезно предоставлены Ласло Макрай (Университет ветеринарной медицины, Будапешт). Shigella flexneri M90T любезно предоставил Zoltán Tigyi (Университет Печа). Энтероинвазивные штаммы E. coli (EIEC) и Shigella (за исключением S. sonnei 866-F) происходят из Венгерской национальной коллекции медицинских бактерий (HNCMB), Будапешт.Таблица 2
Специфичность хозяина и эффективность посева двух подгрупп новых Т5-подобных бактериофагов.
245 6Strain Pathotype/serovar /species Serogroup or serotype Phage chee24 Phage chee130_1 Strain reference II95 -36 EIEC O121 − − данное исследование 20 EIEC − − this study Bra2 26 EIEC O152 +++ +++ this study Saigon EIEC O164 − − this study E2348/69 EPEC O127:H6 − + Iguchi et al. , 2009536 UPEC O6:K15:h41 − − Hochhut et al., 2006 IHE3034 ExPEC O18:K1:H7 − − Moriel et al., 2010 E250 APEC O1:K1:H7 − − Tóth et al. , 2003T22 atypical O157:h53 − − Tóth et al., 2009 Е22 EHEC O103: H3 ++ — Marchès et al., 2003 Sakai EHEC 57: 29505957 4057 4057 4057 4057 4057 4057. EDL933 EHEC O157:H7 − − Perna et al., 2001 ICC169 Citrobacter rodentium N/A − − Wiles et al., 2005 20080 Shigella dysenteriae 1A N/A +++ − this study M90T Shigella flexner i N/A − − this study 20038 Shigella boydii N/A − − this study 866-F Shigella sonnei N/A +++ +++ Allué-Guardia et al. , 201120045 Shigella sonnei N/ A +++ − this study 1201 Salmonella Typhimurium 1 N/A +++ − this study 1202 Сальмонелла Infantis N/A + − this study 1203 Salmonella Panama N/A +++ − this study 1199 Salmonella Typhi N/A − − this study 1198 Salmonella Gallinarum N/A − − this study 1200 Salmonella Enteritidis N/A − − this study 5871 * ESBL E. coli O15 − − this study 18531 ** ESBL E. coli O73 ++ + this study 29095 ** ESBL E. Coli O90 — — 0. «text»:»h20407″,»term_id»:»875229″,»term_text»:»h20407″}}h20407ETEC O78:h21:K80 + + Evans 1 et al. MG1655 E. coli K-12 O16:h58 +++ +++ Blattner et al., 1997 Открыто в отдельном окне
+: от 1 до 10 −3 ; ++: между 10 −4 −10 −6 ; +: 10 −7 и ниже; −: без лизиса .
* Устойчивость к БЛРС, ципрофлоксацину, гентамицину, тетрациклину .
** Устойчивость к ESBL, ципрофлоксацину, гентамицину, сульфаметоксазолу/триметоприму .
Штаммы E. coli , устойчивые к БЛРС, были выделены из внекишечных инфекций человека, штамм Shigella flexneri был выделен из фекалий человека, а штаммы Salmonella были выделены из пищевых продуктов или животных. Эти изоляты были идентифицированы биохимически и серотипированы с использованием О-специфических иммунных сывороток (данные не показаны).
Эффективность посева (ЭОП) определяли путем применения серийных разведений суспензий бактериофагов в точечных анализах. Отношение титра бактериофага на каждом штамме к титру, измеренному на E. coli MG1655, считали ЭОП бактериофага на данном штамме.
Одноэтапные эксперименты по выращиванию
Одноэтапный эксперимент по выращиванию для определения размера взрыва был проведен на репрезентативных бактериофагах chee24 и chee130_1 на E.
coli MG1655 в соответствии с описанием E. Bassiri 1 с некоторыми изменениями. Вкратце, 5 × 10 8 бактерий смешивали с 5 × 10 6 бактериофагов в бульоне Луриа (LB), устанавливая множественность инфекции (MOI) на 0,01. Смесь инкубировали 10 мин при комнатной температуре для начальной адсорбции, затем разбавляли в 10 4 раза и в общем объеме 10 мл инкубировали при 37°С в течение 1 ч. Аликвоты по 100 мкл отбирали каждые 5 мин и высевали на слоистый мягкий агар для подсчета. Эксперименты проводились в три независимых момента времени по две параллели каждый. Размер всплеска определяли как отношение среднего количества бактериофагов на исходном уровне и среднего количества бактериофагов после всплеска.Испытания на термостабильность
Термостабильность бактериофагов тестировали следующим образом: 1 мл раствора, содержащего 100-кратное разведение исходного раствора бактериофага в LB, инкубировали в течение 1 ч при 25°C, 37°C, 42°C, и 80°С.
После инкубации титр обработанных запасов бактериофагов определяли с помощью точечного анализа на E. coli MG1655. Опыты проводились в двух параллелях при каждой температуре.Тесты на переносимость pH
Стабильность бактериофагов при различных значениях pH проверяли путем инкубации 100-кратных разведений бактериофагов в течение 1 ч при 37°C в 1 мл LB с доведением pH до 3, 5, 7 и 9растворами HCl и NaOH. После инкубации титр обработанных запасов бактериофагов определяли с помощью точечного анализа на E. coli MG1655. Опыты проводили в двух параллелях при каждом значении рН.
Определение морфологии бактериофагов
Бактериофаги исследовали методом просвечивающей электронной микроскопии (ПЭМ). Вкратце, капли суспензии бактериофагов с высоким титром помещали на парафильм, абсорбировали на углеродную пленку, промывали буфером ТЕ (10 мМ ТРИС, 1 мМ ЭДТА, рН 6,9).) и отрицательно окрашены 2% водным раствором уранилацетата, рН 5,0. Углеродную пленку собирали с помощью медных сеток 300 меш, негативное пятно доступа удаляли фильтровальной бумагой и затем сушили на воздухе.
Образцы исследовали в просвечивающем электронном микроскопе TEM 910 (Carl Zeiss, Oberkochen) при ускоряющем напряжении 80 кВ. Изображения записывали в цифровом виде с калиброванным увеличением с помощью ПЗС-камеры с медленным сканированием (ProScan, 1024 × 1024, Scheuring, Германия) с программным обеспечением ITEM (Olympus Soft Imaging Solutions, Мюнстер, Германия). Контрастность и яркость настраивались с помощью Adobe Photoshop CS3.Выделение ДНК бактериофага
ДНК бактериофага выделяли из запасов бактериофага с концентрацией не менее 10 11 БОЕ/мл. Фенол-хлороформный метод, описанный Sambrook et al. (1987) использовали для выделения с модификациями, описанными Tóth et al. (2016). Вкратце, для удаления ДНК и РНК, не связанных с бактериофагами, из 700 мкл суспензии бактериофага образец обрабатывали ДНКазой I и РНКазой A степени амплификации (Sigma Aldrich) в конечной концентрации 10 мкг/мл в течение 30 мин при 37°C. Для лизиса бактериофагом протеиназу К (Sigma Aldrich) в конечной концентрации 6,6 мкг/мл добавляли к лизирующему буферу (50 мМ Трис-HCl (рН 8,0), 10 мМ ЭДТА, 0,5% ДСН) и инкубировали при 65°С в течение 30 мин.
Суспензию охлаждали до комнатной температуры и смешивали с 750 мкл смеси 1:1 уравновешенного фенола и хлороформа. После 5 мин инкубации при комнатной температуре белки удаляли центрифугированием при 10000×g в течение 5 мин. Водную фазу переносили в чистую пробирку Эппендорфа и осаждали ДНК бактериофага после добавления 0,1 объема 3 М ацетата калия, рН 5,5 с 0,7 объема изопропанола на льду в течение 20 мин. ДНК собирали центрифугированием при 13000×g в течение 10 мин, промывали 600 мкл 70%-ного этанола и растворяли в 30 мкл буфера 10 мМ Трис-HCl, 1 мМ ЭДТА, рН 8,0.Секвенирование и анализ генома бактериофага
Библиотеки секвенирования геномной ДНК готовили с использованием набора Nextera XT (Illumina, Eindhoven, NL). Секвенирование проводили с использованием набора реагентов Nextseq Mid-output v2 (2 × 150 п.н.) на Illumina NextSeq 500. Сборку проводили с помощью SPAdes (Банкевич и др., 2012). Геном был аннотирован с помощью сервера RAST (Overbeek et al., 2014). Для филогенетического анализа использовали гены orf5c (T5.
035) и ДНК-полимеразы бактериофага (T5.122). Последовательности ДНК этих двух генов из всех бактериофагов, определенных в этом исследовании, и эталонных геномов T5 (дополнительная таблица 1) были выровнены с использованием Clustal Omega (Sievers et al., 2011). Филогенетическое дерево было визуализировано с помощью MEGA5.2 (Hall, 2013). Для филогении последовательностей целых бактериофагов использовалась программа VICTOR (Meier-Kolthoff and Goeker, 2017) с настройками по умолчанию и всеми эталонными геномами T5 (дополнительная таблица 1). Определение гомологии последовательностей и идентификацию SNP проводили с использованием инструментов BLAST, доступных на веб-сайте NCBI. Выравнивание генома визуализировали с помощью Easyfig (Sullivan et al., 2011) и модифицировали с помощью Inkscape. Левый и правый повторы были определены с помощью анализа накопления необработанных считываний секвенирования после картирования с помощью CLC workbench 9..0 (Qiagen, Хильден, Германия). Повторы были определены как > 130% среднего охвата в конкретном месте (на 30% больше охвата, чем средний охват).Номер доступа последовательности
Полногеномные последовательности всех бактериофагов, определенные в данном исследовании, были депонированы в базе данных GenBank под номерами доступа. {«type»:»entrez-нуклеотид»,»attrs»:{«text»:»MF431730″,»term_id»:»1345606605″,»term_text»:»MF431730″}}MF431730- {«type»:»entrez -нуклеотид»,»attrs»:{«текст»:»MF431741″,»term_id»:»1345606759″,»term_text»:»MF431741″}}MF431741.
Выделение бактериофагов
Используя производных штаммов E. coli K-12 MG1655 и C600 в качестве индикатора и штамма для размножения, мы выделили 12 новых бактериофагов из различных образцов пищевых продуктов. . Происхождение, а также подгруппы бактериофагов приведены в таблице . Бактериофаги были выделены либо из мяса, мясных продуктов (например, колбас) или сыра
Морфология
Все новые выделенные бактериофаги имели морфологию Siphoviridae, с средняя длина головы ~ 85 нм, ширина головы ~ 75 нм и гибкий несокращающийся хвост длиной ~ 200 нм.
В качестве представителя показана морфология T5-подобного бактериофага chee24 (рис. 9).0014Открыть в отдельном окне
Трансмиссионная электронная микрофотография бактериофага 24, показывающая морфологию Siphoviridae.
Спектр хозяев, эффективность посева, размер пакета и стабильность
Чтобы оценить специфичность бактериофагов к хозяину, мы протестировали их литическую способность на большом количестве патогенных энтеробактерий и клинических штаммов БЛРС с множественной лекарственной устойчивостью, перечисленных в таблице. Мы обнаружили, что новые T5-подобные бактериофаги демонстрируют два различных паттерна лизиса, которые мы обозначаем репрезентативным бактериофагом каждого: бактериофаг chee24 (группа chee24) и бактериофаг chee130_1 (группа chee130_1). Группировка бактериофагов указана в табл. Значения EOP для этих двух бактериофагов определяли точечным анализом. Спектр лизиса и значения ЭОП приведены в табл.
Помимо производных штаммов К-12, несколько штаммов E.
coli, Shigella и Salmonella , представляющих разные патотипы и серовары, оказались чувствительными к бактериофагам. В то время как бактериофаг chee130_1 лизировал энтеропатогенные штаммы E. coli (EPEC), EIEC и Shigella sonnei , бактериофаг 24 индуцировал лизис сероваров Salmonella , штаммов EHEC O103:h3 и Shigella dysenteriae 90. Оба бактериофага показали лизис на энтеротоксигенных Тип штамма E. coli (ETEC) {«type»:»entrez-нуклеотид»,»attrs»:{«text»:»h20407″,»term_id»:»875229″,»term_text»:»h20407″} }h20407. Однако следует отметить, что в каждом случае значения ЭОП были на порядки ниже по сравнению со штаммами К-12, а у некоторых штаммов дикого типа морфология бляшек была непрозрачной, в отличие от наблюдаемых прозрачных бляшек диаметром ~2 мм. со штаммом MG1655.Размер всплеска определяли в экспериментах по выращиванию в один этап на E. coli MG1655 (рис. ). Величина всплеска бактериофагов chee24 и chee130_1 оказалась разной, в случае первого она составила около 1000 БОЕ/клетку, а в случае chee130_1 размер всплеска составил около 44 БОЕ/клетку.
В обоих случаях всплеск произошел через ~45 мин после окончания начального поглощения.Открыть в отдельном окне
Одношаговые кривые роста новых T5-подобных бактериофагов на штамме E. coli MG1655. Треугольниками отмечены значения БОЕ бактериофага chee24, квадратами – бактериофага chee130_1. Планки погрешностей указывают на стандартные отклонения от результатов трех независимых трехкратных экспериментов.
Эксперименты по термостабильности показали, что инкубация 10 10 БОЕ запасов бактериофагов при 25°C, 37°C и 42°C в течение 1 часа не оказывала существенного влияния на EOP бактериофагов (данные не показаны). Однако инкубация при 80°C вызывала 10 4 — среднекратное снижение титра бактериофагов.
Значение рН 3 полностью дезактивировало бактериофаги, так как бляшек не наблюдалось даже в местах концентрированных суспензий. С другой стороны, не наблюдалось значительного снижения титра при значениях pH 5, 7 и 9 после 1 ч инкубации (данные не представлены).
Анализ последовательности всего генома
Для изучения геномных структур и потенциального разнообразия геномов были определены последовательности геномов всех 12 вновь выделенных бактериофагов и проведено подробное геномное сравнение. Секвенирование генома бактериофагов показало, что все бактериофаги имеют геном линейной двухцепочечной ДНК и относятся к Т5-подобным бактериофагам. Две подгруппы с различной специфичностью хозяина также имеют незначительные генетические различия. Геном бактериофагов «группы сыр24» составляет 120618 (бактериофаг свиной27), 120620 (бактериофаг саус111К), 120622 (бактериофаг сыр24, свиной29и saus47N) или 120 629 п.н. (бактериофаг poul124) и содержит 165 (saus47N, saus111K, poul124), 166 (chee24, свинина29) или 168 (свинина27) белок-кодирующих CDS, а также 18 генов тРНК соответственно. Его содержание GC составляет 39,3%. С другой стороны, размер генома бактериофагов «группы chee130_1» составляет 121 986 п.н., с 164 (saus132, poul149), 165 (cott162) или 167 (chee130_1, chee158, saus176N) белок-кодирующими CDS, 20 генами тРНК и содержание GC 39,7%.
Эти группы соответствовали различному спектру хозяев, наблюдаемому у бактериофагов, что указывает на то, что chee24-подобные и chee130_1-подобные бактериофаги образуют разные генетические группы. Список ОРС в сравнении друг с другом и с исходным бактериофагом Т5 (GenBank {«type»:»entrez-нуклеотид»,»attrs»:{«text»:»AY543070.1″,»term_id»:»45774915″,»term_text»:»AY543070.1″}}AY543070.1) показаны в дополнительных таблицах 2, 3. Члены обеих подгрупп показали характерные функциональные области в своих геномах, что является общей чертой Т5-подобных фагов. , имеющие предранние, ранние и поздние участки генома, названные в честь времени их транскрипции, а также участок терминального повтора на 5′- и 3′-конце, длина которого у chee24-подобных фагов составляет 9968 п.н. фагов группы chee130_1.Для бактериофагов группы chee24 бактериофаг свинина27 отличался на 1 SNP в терминальном повторе, тогда как для фагов группы chee130_1 все терминальные повторы фагов, отличных от chee130_1, отличались от присутствующих в прототипе, с saus132, chee158, cott162 и saus176N имеющий 3 SNP, и poul149содержит 4 SNP.На уровне нуклеотидов недавно идентифицированные Т5-подобные бактериофаги не только имеют одинаковый размер генома в пределах двух подгрупп — группы chee24 и группы chee130_1-, но и при сравнении с «типовым бактериофагом» их групп в качестве эталонного бактериофага chee24 или chee130_1, соответственно, отличаются всего 2–5 позициями нуклеотидов. Следует отметить, что в случае бактериофагов pig27 и saus47N один SNP вызывает преждевременный стоп-кодон в генах белка бактериофага с неизвестной функцией. В случае бактериофагов saus132, poul149и chee158 в гене хвостового отростка бактериофага имеются преждевременные стоп-кодоны. Краткое изложение этих геномных местоположений приведено в таблице.
Таблица 3
Однонуклеотидные полиморфизмы, показанные T5-подобными бактериофагами из подгруппы, подобной chee24 (A) и подгруппы, подобной chee130_1 (B), по сравнению с типовым бактериофагом каждой подгруппы.
90 Фаговый белок00057397474 9000974743974743974749705656777. 0742 TА Позиция в фаге 24 ORF Predicted function chee24 pork27 pork29 saus47N saus111K * poul124 ** Замена аминокислот 24634 42 Белок фага С А А 0005 C A A V65L 57794 101 Phage protein G G G A G G W22 * 84550 129 Phage tail fibers C C C T T T G700D 84602 129 Phage tail fibers T G G G G G T683P 86896 130 Phage protein C A C C C C E59 * B Position in phage chee130_1 ORF Predicted function chee130_1 saus132 poul149 chee158 cott162 saus176N Замена аминокислот 595 2 G A A G G G synonymous 1774 4 Probable A1 protein C C C G C C G493A 1919 4 Probable A1 protein G G G G A G L445F 2501 4 Probable A1 protein A A G A A A synonymous 2707 4 Probable A1 protein G G G G G A R182W 3301 intergenic region A C C C C C N/A 3303 intergenic region T A a A A A N/A N/A N/A N/A N/A C C T T T N/A 86944 133 Phage tail fibers G G G A G G Q210 * 86950 133 Phage tail fibers C C C C T C A212T 87643 133 Phage tail fibers G A A G G G Q443 * Открыть в отдельном окне
* В позиции 101429-101430 удалено АА .
** В позиции 101429-101430 АА удален. В начале вставляется ACT, а в конце вставляется CGTG .
Буквы, выделенные жирным шрифтом, относятся к нуклеотидам, отличным от найденных в фагах chee24 и chee130_1 в тех же позициях соответственно .
Ни один из новых охарактеризованных геномов бактериофагов не содержал каких-либо известных генов вирулентности, токсинов, генов, кодирующих устойчивость к антибиотикам, и генов-регуляторов вирулентности.
Филогенетический анализ
Для выявления филогенетического положения бактериофагов chee24 и chee130_1 среди Т5-подобных фагов было проведено несколько сравнений. Сравнение на основе генов orf5c (T5.035) и ДНК-полимеразы бактериофага (T5.122) бактериофага T5 ({«type»:»entrez-нуклеотид»,»attrs»:{«text»:»AY6.1″,»term_id»:»51511954″,»term_text»:»AY6.1″}}AY6.1) показали, что хотя бактериофаг 24 оказался ближайшим родственником бактериофагов E. coli DT57C и DT571/2,
Открыть в отдельном окне
Филогенетическое древо Т5-подобных бактериофагов, основанное на выравнивании Clustal Omega генов orf5c (T5.
035, )Открыть в отдельном окне А ) и ДНК-полимераза фага (Т5.122, B ) бактериофага Т5 ({«type»:»entrez-нуклеотид»,»attrs»:{«text»:»AY6.1″,»term_id»:»51511954″,»term_text»:»AY6.1 «}}AY6.1). Шкала представляет процент гомологии. Список включенных геномов T5-подобных фагов с их инвентарными номерами в GenBank приведен в дополнительной таблице 1.
Филогенез всех T5-подобных бактериофагов, основанный на полном геноме, ясно показал, что фаги, выделенные в текущем исследовании, являются членами T5-подобных фагов. как бактериофаги (рис. ). Топология этого дерева полностью отличается от топологии, основанной на отдельных генах, поскольку два репрезентативных фага помещаются рядом с геномами фагов, которые также оказались ближайшими гомологами с помощью анализа BLAST. Выравнивание T5-подобных геномов с взаимными гомологиями, включая представителей двух подгрупп, представлено на рис. , показывающем 5′-конец поздней области как наиболее вариабельный, который содержит в основном гены хвостовых отростков.
Открыть в отдельном окне
Филогенетическое дерево Т5-подобных бактериофагов, полученное с помощью полногеномного анализа VICTOR. Шкала представляет процент гомологии. Список включенных геномов T5-подобных фагов с их инвентарными номерами GenBank приведен в дополнительной таблице 1.
Открыть в отдельном окне
Выравнивание целых геномов T5-подобных бактериофагов. Номера доступа фагов в GenBank приведены в дополнительной таблице 1. Стрелки разных цветов представляют ORF, связанные с областями генома, указанными вверху. Серая полоса в правом нижнем углу показывает процент идентичности, связанный с цветом полос, соединяющих гомологичные области.
При сравнении геномов бактериофагов chee24 и chee130_1 на уровне нуклеотидов области, демонстрирующие низкую гомологию друг другу, представляли собой область хвостового отростка и открытую рамку считывания, кодирующую ген белка, связывающего рецептор, ассоциированный с бактериофагом. Все эти особенности, по-видимому, способствуют различиям, наблюдаемым в специфичности двух групп бактериофагов к хозяину.
На уровне нуклеотидов ближайшим гомологом бактериофага chee24 является бактериофаг SPC35 (номер GenBank {«type»:»entrez-нуклеотид»,»attrs»:{«text»:»HQ406778″,»term_id»:»321272088″,» term_text»:»HQ406778″}}HQ406778.). Кроме того, два гипотетических белка, ген тимидилатсинтазы и две субъединицы эндонуклеазы HNH, белок, связывающий рецептор (CDS {«type»:»entrez-protein»,»attrs»:{«text»:»ADW80123.1 «,»term_id»:»321272231″,»term_text»:»ADW80123.1″}}ADW80123.1) из SPC35 также отсутствует в chee24, что предполагает другую специфичность хоста. Бактериофаг EPS7 (GenBank № {«type»:»entrez-нуклеотид»,»attrs»:{«text»:»CP000917″,»term_id»:»182630511″,»term_text»:»CP000917″}}CP000917) является ближайшим гомологом бактериофага chee130_1 из ранее секвенированных T5-подобных бактериофагов и несет гомологи ко всем рецептор-ассоциированным белкам, хотя сходство не является 100% в каждом случае.Кроме того, у chee130_1 отсутствуют несколько гипотетических белков, а также предполагаемый хвостовой белок AGC_0146. Гены с низкой гомологией и гены, отсутствующие у бактериофагов подгруппы chee24 или chee130_1, перечислены в таблице 9.0014Таблица 4
Список генов и областей с низкой гомологией или отсутствующих у chee24-подобных (A) и chee130_1-подобных (B) бактериофагов по сравнению с их ближайшими гомологами-бактериофагами.
905″: entrez-protein»,»attrs»:{«text»:»ADW80013.1″,»term_id»:»321272121″,»term_text»:»ADW80013.1″}}ADW80013.1 60A Phage chee24 Phage SPC35 ORF Predicted function CDS Предсказанная функция нет межгенная область {«type»:»entrez-protein»,»attrs»:{«text»:»ADW79993. 1″,»term_id»:»321272101″,» term_text»:»ADW79993.1″}}ADW79993.1гипотетический белок нет межгенная область {«type»:»entrez-protein»,»attrs»:{«text»:»ADW79995 .1″,»term_id»:»321272103″,»term_text»:»ADW79995.1″}}ADW79995.1 гипотетический белок 7 фаговый белок {«type»:»entrez-protein»,»attrs»:{«text»:»ADW79996.1″,»term_id»:»321272104″,»term_text»:»ADW79996.1″} }ADW79996.1 гипотетический белок 7 фаговый белок {«type»:»entrez-protein»,»attrs»:{«text»:»ADW79997. 1″,»term_id»:» 321272105″,»term_text»:»ADW79997.1″}}ADW79997.1гипотетический белок нет межгенная область {«type»:»entrez-protein»,»attrs»:{«type»:»entrez-protein»,»attrs» «:»ADW80009.1″,»term_id»:»321272117″,»term_text»:»ADW80009.1″}}ADW80009.1 гипотетический белок 31 серин-треонинпротеинфосфатаза серин-треонинпротеинфосфатаза 41 фаговый белок {«type»:»entrez-protein»,»attrs»:{«text»:»ADW80023. 1″,»term_id»:»321272131″,»term_text»:»ADW80023 .1″}}ADW80023.1гипотетический белок 41 фаговый белок {«type»:»entrez-protein»,»attrs»:{«text»:»ADW80025.1″,»term_id»:»321272133″,» term_text»:»ADW80025.1″}}ADW80025.1 гипотетический белок нет межгенная область {«type»:»entrez-protein»,»attrs»:{«text»:»ADW80038 .1″,»term_id»:»321272146″,»term_text»:»ADW80038.1″}}ADW80038.1 гипотетический белок нет межгенная область {«type»:»entrez-protein»,»attrs»:{«text»:»ADW80040. 1″,»term_id»:»321272148″,»term_text»:»ADW80040.1″} }ADW80040.1гипотетический белок 91 тимидилатсинтаза {«type»:»entrez-protein»,»attrs»:{«text»:»ADW80064.1″,»term_id»:» 321272172″,»term_text»:»ADW80064.1″}}ADW80064.1 тимидилатсинтаза нет межгенная область {«type»:»entrez-protein»,»attrs «:»ADW80067.1″,»term_id»:»321272175″,»term_text»:»ADW80067.1″}}ADW80067.1 предполагаемая H-N-H-эндонуклеаза P-TflVIII 99 фаговый белок {«type»:»entrez-protein»,»attrs»:{«text»:»ADW80071. 1″,»term_id»: «321272179»,»term_text»:»ADW80071.1″}}ADW80071.1гипотетический белок нет межгенная область {«type»:»entrez-protein»,»attrs»:{» text»:»ADW80078.1″,»term_id»:»321272186″,»term_text»:»ADW80078.1″}}ADW80078.1 гипотетический белок 134 белок-рулетка с длиной хвоста фага {«type»:»entrez-protein»,»attrs»:{«text»:»ADW80108.1″,»term_id»:»321272216″,»term_text «:»ADW80108.1»}}ADW80108.1 порообразующий белок кончика хвоста 145 волокна хвоста фага {«type»:»entrez-protein»,»attrs»:{«text «:»ADW80118. 1″,»term_id»:»321272226″,»term_text»:»ADW80118.1″}}ADW80118.1предполагаемый хвостовой белок 149 фаг-ассоциированный рецептор-связывающий белок {«type»:»entrez-protein»,»attrs»:{«text»:»ADW80118.1″,»term_id»:»321272226″,»term_text»:»ADW80118.1″}}ADW80118.1 receptor-binding protein B Phage chee130_1 Phage EPS7 ORF Predicted function CDS Predicted function 9 фаговый белок {«type»:»entrez-protein»,»attrs»:{«text»:»ACB97450. 1″,»term_id»:»182630518″,»term_text»:»ACB97450.1″}} ACB97450.1гипотетический белок нет межгенная область {«type»:»entrez-protein»,»attrs»:{«text»:»ACB97451.1″,»term_id»:»182630519 «,»term_text»:»ACB97451.1″}}ACB97451.1 гипотетический белок нет межгенная область {«type»:»entrez-protein»,»attrs»:{«text» :»ACB97452.1″,»term_id»:»182630520″,»term_text»:»ACB97452.1″}}ACB97452.1 гипотетический белок нет межгенная область {«type»:»in»entrez» ,»attrs»:{«text»:»ACB97458. 1″,»term_id»:»182630526″,»term_text»:»ACB97458.1″}}ACB97458.1гипотетический белок нет межгенный регион {«type»:»entrez-protein»,»attrs»:{«text»:»ACB97467.1″,»term_id»:»182630535″,»term_text»:»ACB97467.1″}}ACB97467.1 гипотетический белок 26 фаговый белок {«type»:»entrez-protein»,»attrs»:{«text»:»ACB97475.1″,»term_id»:»182630543″, «term_text»:»ACB97475.1″}}ACB97475.1 гипотетический белок 28 фосфоэстераза {«type»:»entrez-protein»,»attrs»:{«text»:»ACB9747 . 1″,»term_id»:»182630546″,»term_text»:»ACB97478.1″}}ACB97478.1предполагаемая серин/треонинфосфатаза 29 фаговый белок {«type»:»entrez-protein»,»attrs»:{«text»:»ACB97479.1″,»term_id»:»182630547″,»term_text»:»ACB97479 .1″}}ACB97479.1 гипотетический белок 30 серин/треонинфосфатаза {«type»:»entrez-protein»,»attrs»:{«text»:»ACB97480.1″ ,»term_id»:»182630548″,»term_text»:»ACB97480.1″}}ACB97480.1 гипотетический белок 31 ассоциированная с фагом самонаводящаяся эндонуклеаза {«type»:»entrez-protein»,»attrs»:{«text»:»ACB97482. 1″,»term_id»:»182630550″,»term_text»:»ACB97482.1″}}ACB97482.1гипотетический белок нет межгенная область {«type»:»entrez-protein»,»attrs»:{«text»:»ACB97510.1″,»term_id»:»182630578″,» term_text»:»ACB97510.1″}}ACB97510.1 гипотетический белок нет межгенная область {«type»:»entrez-protein»,»attrs»:{«text»:»ACB97513.1″,»term_id»:»182630581″,»term_text»:»ACB97513.1″}}ACB97513.1 ро-ассоциированная, спирально-спиральная, содержащая протеинкиназу 2 60 0-фаговый белок «type»:»entrez-protein»,»attrs»:{«text»:»ACB97515. 1″,»term_id»:»182630583″,»term_text»:»ACB97515.1″}}ACB97515.1гипотетический белок 61 гипотетический белок {«type»:»entrez-protein»,»attrs»:{«text»:»ACB97516.1″,»term_id»:»182630584″,»term_text»: «ACB97516.1»}}ACB97516.1 гипотетический белок нет межгенная область {«type»:»entrez-protein»,»attrs»:{«text»:»ACB97518.1″,»term_id «:»182630586″,»term_text»:»ACB97518.1″}}ACB97518.1 гипотетический белок 65 фаговый белок {«type»:»entrez-protein»,»attrs-protein»,»attrs-protein» {«text»:»ACB97519. 1″,»term_id»:»182630587″,»term_text»:»ACB97519.1″}}ACB97519.1гипотетический белок 101 неизвестно {«type»:»entrez-protein»,»attrs»:{«text»:»ACB97556.1″,»term_id»:»182630624″,»term_text»:»ACB97556. 1″}}ACB97556.1 гипотетический белок 105 НАД-зависимая протеиндеацетилаза семейства SIR2 {«type»:»entrez-protein»,»attrs»:{«text»:»ACB97560 .1″,»term_id»:»182630628″,»term_text»:»ACB97560.1″}}ACB97560.1 предполагаемый Sir2-подобный белок 109 гипотетический белок {«type»:»entrez-protein»,»attrs»:{«text»:»ACB97569. 1″,»term_id»:»182630637″,»term_text»:»ACB97569.1″}} ACB97569.1гипотетический белок 134 хвостовые волокна фага {«type»:»entrez-protein»,»attrs»:{«text»:»ACB97589.1″,»term_id»:» 182630657″,»term_text»:»ACB97589.1″}}ACB97589.1 предполагаемый белок хвоста фага 139 белок хвостовой рулетки фага {«type»:»entrez-protein»,»attrs»:{«text»:»ACB97594.1″,»term_id»:»182630662″,»term_text»:»ACB97594.1″}}ACB97594.1 порообразующий белок кончика хвоста 139 фаговый белок-рулетка длины хвоста {«type»:»entrez-protein»,»attrs»:{«text»:»ACB97595. 1″,» term_id»:»182630663″,»term_text»:»ACB97595.1″}}ACB97595.1порообразующий белок кончика хвоста pb2 нет межгенная область {«type»:»inentrez-prote «,»атрибуты»:{«текст»:»ACB97613.1″,»term_id»:»182630681″,»term_text»:»ACB97613.1″}}ACB97613.1 гипотетический белок Открыть в отдельном окне
подгруппа эталонного бактериофага — SPC35 (GenBank {«type»:»entrez-нуклеотид»,»attrs»:{«text»:»HQ406778″,»term_id»:»321272088″,»term_text»:»HQ406778″}}HQ406778 ), в случае chee130_1-подобных бактериофагов это EPS7 (GenBank {«type»:»entrez-нуклеотид»,»attrs»:{«text»:»CP000917″,»term_id»:»182630511″,»term_text» :»CP000917″}}CP000917). Соответствующие, хотя в некоторых случаях негомологичные ORF или межгенные области отображаются в том же ряду .
12 новых Т5-подобных бактериофагов из независимых образцов пищевых продуктов различного животного происхождения со всего мира Эти новые бактериофаги представляли две подгруппы в зависимости от их специфичности в отношении хозяина, в то время как наиболее эффективными хозяевами бактериофагов были производные К-12 E. coli , несколько штаммов, представляющих важные кишечные патотипы E. coli, Shigella и Salmonella , также оказались чувствительными к бактериофагам. Морфология Siphoviridae и геномные характеристики, включая размер генома, архитектуру генома, низкое содержание GC и гомологии последовательностей, показали, что бактериофаги являются T5-подобными бактериофагами.Важной особенностью этих вновь обнаруженных фагов является то, что они были выделены из образцов пищевых продуктов, в отличие от ранее выделенных Т5-подобных бактериофагов, происходящих из сточных и фекальных источников (Kim, Ryu, 2011; Голомидова и др., 2015). Также имеется один случай, когда T5-подобный фаг был выделен и охарактеризован из коммерчески доступного коктейля бактериофагов, разработанного против кишечных патогенов (Piya et al.
, 2015). Наши результаты показывают, что пища может быть богатым источником бактериофагов, эффективных против патогенных бактерий, потенциально обитающих в той же среде. Эти фаги сохраняют свою инфекционную способность в этой среде, что позволяет предположить, что фаги пищевого происхождения являются многообещающими кандидатами для биоконтроля против бактерий пищевого происхождения.Наши результаты также показывают огромное разнообразие бактериофагов, даже из хорошо охарактеризованных групп, таких как T5-подобные бактериофаги, поскольку подгруппы chee24 и chee130_1 отличаются от ближайших родственных бактериофагов. Наши данные показывают, что наиболее важные различия находятся внутри генов, кодирующих белки, связывающие рецепторы (таблица, рисунок). Ближайшим колинеарным родственником бактериофага chee24 является SPC35, который, как сообщается, лизирует штаммов Salmonella Typhimurium и Enteritidis, а также большой набор производных K-12 и комменсалов E. coli (Ким и Рю, 2011 г.
). Однако спектр хозяев фагов группы chee24 особенно широк и включает штамм, представляющий один из так называемых «большой шестерки» серотипов EHEC (O103:h3), а также штамм ESBL E. coli серогруппы O73, выделенный из внекишечной инфекции. Различия в ORF 145 и 149, аннотированных как «волокна хвоста фага» и «белок, связывающий рецептор», могут объяснить несоответствие.Мы хотели проверить эффективность новых T5-подобных фагов на целевых штаммах и одношаговый рост на E. coli MG1655 давала размеры взрывов, сравнимые с размерами T5-подобного бактериофага phiLLS (Amarillas et al., 2017).
Ближайший гомолог подгруппы chee130_1, EPS7, способен лизировать те же серовары Salmonella и семь безымянных штаммов E. coli (Hong et al., 2008), однако фаг chee130_1 не лизировал штаммов Salmonella8 . С другой стороны, фаг chee130_1 был способен размножаться на штаммах Shigella sonnei , EIEC и EPEC. Все эти различия в диапазоне хозяев подчеркивают потенциальную роль, которую могут играть гены, проявляющие низкую гомологию с близкородственными бактериофагами (таблица), в специфичности хозяина.
В случае chee130_1, ORF134, которая аннотирована как «волокна хвоста фага», имеет очень низкую гомологию с соответствующим геном в EPS7 и, по-видимому, поддерживает это мнение. Однако интересно отметить, что в ORF133, еще одном гене, аннотированном как волокно хвоста фага, имеются преждевременные стоп-кодоны в случае бактериофагов saus132, poul149.и чи158. Тем не менее, эти мутации, по-видимому, не оказывали никакого влияния на специфичность этих бактериофагов к бактериальному хозяину по сравнению с другими членами их подгруппы (данные не показаны).Ли и др. (2016) предположили, что, помимо коктейлей бактериофагов, решением для одновременного биоконтроля нескольких патогенов пищевого происхождения может быть клонирование нескольких генов, связывающих рецепторы, в один бактериофаг. Для любой цели важно получить знания о новых бактериофагах, способных инфицировать эти патогены, и оценить спектр их хозяев.
Помимо упомянутых выше исследований перспективных фагов-кандидатов для использования в качестве средств биологической борьбы с патогенными бактериями, уже существует несколько коммерчески доступных фаговых коктейлей, направленных на различные энтеробактериальные патогены пищевого происхождения (обзор Abedon, 2017).
Однако, насколько нам известно, за исключением ShigaShield, который направлен против Shigellae (Soffer et al., 2017), и Microgen ColiProteus, направленного против E. coli и Proteus (McCallin et al., 2013), ни один из компонентов фагов не был секвенирован. Чтобы оценить применимость фага либо для биоконтроля, либо для терапии, необходимо обширное знание его генома, чтобы определить, несет ли фаг «нежелательные» гены. Глубокие знания о генах, определяющих специфичность хозяина или влияющих на стабильность фага в различных средах, также являются ценной информацией при выборе бактериофагов для данной терапевтической или биоконтролирующей цели.Как было отмечено Brüssow (2012), разработка реального лечения фаготерапии может занять годы от выделения фага до проведения клинических испытаний. Этот процесс включает, например, очистку (Bourdin et al., 2014b), а также определение специфичности хозяина (Bourdin et al., 2014a) и проверку биологической безопасности фагов либо на животных (Raya et al.
, 2011), либо на животных. или люди (Sarker et al., 2012, 2016). Обширная генетическая база данных фагов, выделенных из разных источников, представляющих разные роды и спектры хозяев, может ускорить процесс выбора фагов для клинических испытаний.Таким образом, мы идентифицировали 12 новых T5-подобных бактериофагов из образцов пищевых продуктов, которые представляют два новых вида в этой группе бактериофагов. Наше исследование до сих пор показывает, что они соответствуют важным критериям, перечисленным Hagens и Loessner (2010) для бактериофагов, которые можно использовать в качестве биоконтроля: определена последовательность их генома, они хорошо размножаются на непатогенном хозяине, у них отсутствует какая-либо вирулентность, регулятор вирулентности, гены устойчивости к токсинам и антибиотикам. Они также кажутся строго литическими и, как было показано, стабильны при температурах хранения и применения, а также в диапазоне значений pH, обычных для большинства пищевых продуктов.
Все вышеупомянутые свойства в сочетании с их пищевым происхождением, а также их широкий и частично перекрывающийся спектр дополнительных хозяев делают новые T5-подобные бактериофаги ценными кандидатами в качестве эффективных средств биологической борьбы с патогенными энтеробактериями пищевого происхождения.
IT и TC задумали и разработали исследование. DS, LF, MR, JS и IT провели фенотипические эксперименты. LF выполнила секвенирование и предоставила биоинформационные инструменты. DS, LF, TC и IT проанализировали результаты и составили рукопись. Все авторы рассмотрели рукопись.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Эта работа была поддержана проектом PROMISE 7-й рамочной программы ЕС (проект № 265877), Венгерским национальным бюро исследований, разработок и инноваций (NKFIH, грант 124335) и грантами Bundesministerium fuer Bildung und Forschung (BMBF, Германия) в Немецком центре инфекционных исследований (DZIF/номер гранта 8032808811 TC).
1 http://www.sas.upenn.edu/LabManuals/biol275/Table_of_Contents_files/13-PhageGrowth.pdf По состоянию на 20 сентября 2017 г.
Дополнительный материал к этой статье можно найти в Интернете по адресу: https ://www.frontiersin.org/articles/10.3389/fmicb.2018.00202/full#supplementary-material
Щелкните здесь, чтобы просмотреть файл с дополнительными данными. (47K, docx)
- Абедон С. Т. (2017). Клиническое применение бактериофагов в качестве антибактериальных «лекарств»: полезность и прецедент. микробиол. Спектр. 5:БАД-0003-2016. 10.1128/microbiolspec.BAD-0003-2016 [PubMed] [CrossRef] [Google Scholar]
- Аллюэ-Гуардиа А., Гарсия-Альхаро К., Муньеса М. (2011). Ген цитолетального растягивающего токсина типа V, кодирующий бактериофаг, индуцированный из неклинических изолятов Escherichia coli . Заразить. Иммун. 79, 3262–3272. 10.1128/IAI.05071-11 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Amarillas L., Rubí-Rangel L., Chaidez C., González-Robles A., Lightbourn-Rojas L., Леон-Феликс Дж. (2017). Выделение и характеристика phiLLS, нового фага с потенциальным агентом биологической борьбы с множественной лекарственной устойчивостью Кишечная палочка . Фронт. микробиол. 8:1355. 10.3389/fmicb.2017.01355 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Anderson M., Sansonetti P.J., Marteyn B.S. (2016). Разнообразие шигелл и меняющийся ландшафт: идеи для двадцать первого века. Фронт. Клетка. Заразить. микробиол. 6:45. 10.3389/fcimb.2016.00045 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
- Банкевич А., Нурк С., Антипов Д., Гуревич А. А., Дворкин М., Куликов А. С. и др.. (2012 ). SPAdes: новый алгоритм сборки генома и его приложения для секвенирования отдельных клеток. Дж. Вычисл. биол. 19, 455–477. 10.1089/cmb.2012.0021 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Bell G., MacLean C. (2017). Поиск «эволюционно-устойчивых» антибиотиков. Тенденции микробиол. [Epub перед печатью]. 10.1016/j.tim.2017.11.005 [PubMed] [CrossRef] [Google Scholar]
- Блаттнер Ф. Р., Планкетт Г., Блох К. А., Перна Н. Т., Берланд В., Райли М. и др. (1997). Полная последовательность генома Escherichia coli K-12. Наука
277, 1453–1462. 10.1126/science.277.5331.1453 [PubMed] [CrossRef] [Google Scholar]
- Бурден Г., Наварро А., Саркер С. А., Питте А.-К., Кадри Ф., Султана С. и др. (2014a). Охват ассоциированных с диареей изолятов Escherichia coli различного происхождения двумя типами фаговых коктейлей. микроб. Биотехнолог. 7, 165–176. 10.1111/1751-7915.12113 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Бурден Г., Шмитт Б., Марвин Гай Л., Джермонд Дж.-Э., Зубер С., Мишо Л., и др.. (2014b). Амплификация и очистка T4-подобных Escherichia coli фагов для фаговой терапии: от лабораторных до экспериментальных. заявл. Окружающая среда. микробиол. 80, 1469–1476. 10.1128/AEM.03357-13 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Brüssow H. (2012). Что необходимо для того, чтобы фаговая терапия стала реальностью в западной медицине? Вирусология 434, 138–142. 10.1016/j.virol.2012.09.015 [PubMed] [CrossRef] [Google Scholar]
- Carter C. D., Parks A., Abuladze T., Li M., Woolston J., Magnone J., et al. (2012). Коктейль из бактериофагов значительно снижает Escherichia coli O157: Заражение H7 салата и говядины, но не защищает от повторного заражения. Бактериофаг
2, 178–185. 10.4161/bact.22825 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Croxen M.A., Law R.J., Scholz R., Keeney K.M., Wlodarska M., Finlay B.B. (2013). Последние достижения в понимании кишечной патогенности Escherichia coli . клин. микробиол. Откр. 26, 822–880. 10.1128/CMR.00022-13 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Дэвисон Дж. (2015). Предранние функции бактериофага Т5 и его родственников. Бактериофаг 5:e1086500. 10.1080/21597081.2015.1086500 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Эванс Д. Г., Эванс Д. Дж., Пирс Н. Ф. (1973). Различия в реакции тонкого кишечника кроликов на термолабильные и термостабильные энтеротоксины Escherichia coli . Заразить. Иммун. 7, 873–880. [PMC free article] [PubMed] [Google Scholar]
- Голомидова А. К., Куликов Е. Е., Прохоров Н. С., Герреро-Феррейра Р. К., Ксензенко В. Н., Тарасян К. К. и др.. (2015). Полные последовательности генома T5-связанных Бактериофаги Escherichia coli DT57C и DT571/2, выделенные из фекалий лошадей. Арка Вирол. 160, 3133–3137. 10.1007/s00705-015-2582-0 [PubMed] [CrossRef] [Google Scholar]
- Hagens S., Loessner MJ (2010). Бактериофаг для биоконтроля пищевых патогенов: расчеты и соображения. Курс. фарм. Биотехнолог. 11, 58–68. 10.2174/13807429 [PubMed] [CrossRef] [Google Scholar]
- Hall BG (2013). Построение филогенетических деревьев из молекулярных данных с помощью MEGA. Мол. биол. Эвол. 30, 1229–1235. 10.1093/molbev/mst012 [PubMed] [CrossRef] [Google Scholar]
- Хаяши Т., Макино К., Ониши М., Курокава К., Исии К., Йокояма К. и др. (2001). Полная последовательность генома энтерогеморрагической Escherichia coli O157:H7 и сравнение генома с лабораторным штаммом К-12. Рез. ДНК
8, 11–22. 10.1093/dnares/8.1.11 [PubMed] [CrossRef] [Google Scholar]
- Hochhut B., Wilde C., Balling G., Middendorf B., Dobrindt U., Brzuszkiewicz E., et al.. (2006) . Роль остров-ассоциированных интеграз патогенности в пластичности генома уропатогенных Escherichia coli штамм 536. Мол. микробиол. 61, 584–595. 10.1111/j.1365-2958.2006.05255.x [PubMed] [CrossRef] [Google Scholar]
- Хонг Дж., Ким К.-П., Хеу С., Ли С.Дж., Адхья С., Рю С. (2008 г.) ). Идентификация рецептора-хозяина и рецептор-связывающего модуля недавно секвенированного Т5-подобного фага EPS7. ФЭМС микробиол. лат. 289, 202–209. 10.1111/j.1574-6968.2008.01397.x [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Hong Y., Pan Y., Harman NJ, Ebner P.D. (2014). Полные геномные последовательности двух Escherichia coli Фаги O157:H7 эффективны для ограничения загрязнения пищевых продуктов. Объявление генома. 2: e00519–14. 10.1128/genomeA.00519-14 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Hunter P. (2012). Что дальше для антибиотиков?: иммунная система и природа патогенности дают жизненно важные подсказки в борьбе с устойчивыми к антибиотикам бактериями. EMBO Rep. 13, 680–683. 10.1038/embor.2012.101 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Игучи А., Томсон Н. Р., Огура Ю., Сондерс Д., Оока Т., Хендерсон И. Р. и др. (2009). Полная секвенция генома и сравнительный анализ генома энтеропатогенного Escherichia coli O127:H6 штамма E2348/69. Дж. Бактериол. 191, 347–354. 10.1128/JB.01238-08 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Johnson R.P., Gyles C.L., Huff W.E., Ojha S., Huff G.R., Rath N.C., et al.. (2008) . Бактериофаги для профилактики и лечения крупного рогатого скота, птицы и свиней. Аним. Здоровье Рез. Откр. 9, 201–215. 10.1017/S1466252308001576 [PubMed] [CrossRef] [Google Scholar]
- Ким М., Рю С. (2011). Характеристика Т5-подобного колифага, SPC35, и дифференцированного развития устойчивости к SPC35 у Salmonella enterica серовара typhimurium и Escherichia coli . заявл. Окружающая среда. микробиол. 77, 2042–2050 гг. 10.1128/AEM.02504-10 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Lee H., Ku H.-J., Lee D.-H., Kim Y.-T., Shin Х., Рю С. и др. (2016). Характеристика и геномное исследование нового бактериофага HY01, заражающего оба Escherichia coli O157:H7 и Shigella flexneri : потенциал в качестве агента биоконтроля в пищевых продуктах. ПЛОС ОДИН 11:e0168985. 10.1371/journal.pone.0168985 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Marches O., Ledger T. N., Boury M., Ohara M., Tu X., Goffaux F., et al. (2003). Энтеропатогенные и энтерогеморрагические штаммы Escherichia coli доставляют новый эффектор под названием Cif, который блокирует переход G2/M клеточного цикла. Мол. микробиол. 50, 1553–1567. 10.1046/j.1365-2958.2003.03821.x [PubMed] [CrossRef] [Google Scholar]
- McCallin S., Sarker S.A., Barretto C., Sultana S., Berger B., Huq S., et al. (2013). Анализ безопасности российского фагового коктейля: от метагеномного анализа до перорального применения у здоровых людей. Вирусология
443, 187–196. 10.1016/j.virol.2013.05.022 [PubMed] [CrossRef] [Google Scholar]
- Meier-Kolthoff JP, Goeker M. (2017). ВИКТОР: Геномная филогения и классификация прокариотических вирусов. bioRxiv, 107862. 10.1093/bioinformatics/btx440 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Мориэль Д. Г., Бертольди И., Спаньоло А., Марчи С., Розини Р., Неста Б. и др. (2010). Идентификация протективных и ширококонсервативных вакцинных антигенов из генома внекишечной патогенной Escherichia coli . проц. Натл. акад. науч. США 107, 9072–9077. 10.1073/pnas.07107 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Nagy B., Szmolka A., Smole MoŽina S., Kovač J., Strauss A., Schlager S., et al. (2015). Детерминанты вирулентности и устойчивости к противомикробным препаратам веротоксигенных Escherichia coli (VTEC) и полирезистентной E. coli из пищевых продуктов животного происхождения, незаконно ввезенных в ЕС пассажирами авиарейсов. Междунар. Дж. Пищевая микробиология. 209, 52–59. 10.1016/j.ijfoodmicro.2015.06.026 [PubMed] [CrossRef] [Google Scholar]
- Niu Y.D., Stanford K., Kropinski A.M., Ackermann H.-W., Johnson R.P., She Y.-M., et al .. (2012). Геномная, протеомная и физиологическая характеристика T5-подобного бактериофага для борьбы с шига-токсином, продуцирующим Escherichia coli O157:H7. ПЛОС ОДИН 7:e34585. 10.1371/journal.pone.0034585 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Overbeek R., Olson R., Push G.D., Olsen G.J., Davis J.J., Disz T., et al.. ( 2014). SEED и быстрая аннотация микробных геномов с использованием технологии подсистем (RAST). Нуклеиновые Кислоты Res. 42, Д206–Д214. 10.1093/nar/gkt1226 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Перна Н. Т., Планкетт Г., Бурланд В., Мау Б., Гласнер Дж. Д., Роуз Д. Дж. и др.. (2001) . Последовательность генома энтерогеморрагической Escherichia coli O157:H7. Природа
409, 529–533. 10.1038/35054089 [PubMed] [CrossRef] [Google Scholar]
- Пия Д., Се Ю., Эрнандес Моралес А.С., Кути Эверетт Г.Ф. (2015). Полная последовательность генома Salmonella enterica serovar Typhimurium siphophage shivani. Объявление генома. 3: e01443–14. 10.1128/genomeA.01443-14 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Raya R.R., Oot R.A., Moore-Maley B., Wieland S., Callaway T.R., Kutter E.M., et al.. (2011). Природно-резидентные и экзогенно применяемые Т4-подобные и Т5-подобные бактериофаги могут снижать Escherichia coli Уровни O157:H7 в кишках овец. Бактериофаг 1, 15–24. 10.4161/bact.1.1.14175 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Sambrook J., Maniatis T., Fritsch E.F., Laboratory CSH (1987). Молекулярное клонирование: лабораторное руководство, 2-е изд.
Колд-Спринг-Харбор, Нью-Йорк: Лабораторное издательство Колд-Спринг-Харбор; Доступно в Интернете по адресу: http://trove.nla.gov.au/work/13615226 (по состоянию на 1 июня 2017 г. ). [Google Scholar]
- Саркер С. А., Маккаллин С., Барретто С., Бергер Б., Питт А.-С., Султана С. и др. (2012). Пероральное применение Т4-подобного фагового коктейля здоровым взрослым добровольцам из Бангладеш. Вирусология 434, 222–232. 10.1016/ж.вирол.2012.09.002 [PubMed] [CrossRef] [Google Scholar]
- Саркер С. А., Султана С., Рейтелер Г., Мойн Д., Дескомб П., Чартон Ф. и др.. (2016). Пероральная фаготерапия острой бактериальной диареи двумя препаратами колифага: рандомизированное исследование у детей из Бангладеш. ЭБиоМедицина 4, 124–137. 10.1016/j.ebiom.2015.12.023 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Sievers F., Wilm A., Dineen D., Gibson T.J., Karplus K., Li W., et др.. (2011). Быстрое, масштабируемое создание высококачественных множественных выравниваний белков с использованием Clustal Omega. Мол. Сист. биол. 7 539–539. 10.1038/msb.2011.75 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Соффер Н., Вулстон Дж. , Ли М., Дас С., Сулаквелидзе А. (2017). Литический препарат бактериофага для шигелл значительно снижает контаминацию Shigella sonnei в различных пищевых продуктах. ПЛОС ОДИН
12:e0175256. 10.1371/journal.pone.0175256 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Strauch E., Lurz R., Beutin L. (2001). Характеристика бактериофага умеренного пояса 9, кодирующего токсин шига.0457 Шигелла Sonnei . Заразить. Иммун. 69, 7588–7595. 10.1128/IAI.69.12.7588-7595.2001 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Салливан М. Дж., Петти Н. К., Битсон С. А. (2011). Easyfig: визуализатор сравнения геномов. Биоинформатика 27, 1009–1010. 10.1093/bioinformatics/btr039 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Шмолка А., Надь Б. (2013). Комменсал Escherichia coli с множественной лекарственной устойчивостью у животных и его влияние на здоровье населения. Фронт. микробиол. 4:258. 10.3389/fmicb.2013. 00258 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Tóth I., Hérault F., Beutin L., Oswald E. (2003). Продукция цитолетальных растягивающих токсинов патогенными штаммами Escherichia coli , выделенными из источников человека и животных: установление существования нового варианта cdt (тип IV). Дж. Клин. микробиол. 41, 4285–4291. 10.1128/JCM.41.9.4285-4291.2003 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Тот И., Шмидт Х., Кардос Г., Ланц З., Кройцбург К., Дамьянова И., и др.. (2009 г.). Гены вирулентности и молекулярное типирование различных групп штаммов Escherichia coli O157 у крупного рогатого скота. заявл. Окружающая среда. микробиол. 75, 6282–6291. 10.1128/AEM.00873-09 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Тот И., Сваб Д., Балинт Б., Браун-Жак М., Мароти Г. (2016). Сравнительный анализ бактериофага, преобразующего шига-токсин, впервые обнаруженного у Shigella sonnei . Заразить. Жене. Эвол.
37, 150–157. 10.1016/j.meegid.2015.11.022 [PubMed] [CrossRef] [Google Scholar]
- Ван Дж., Цзян Ю., Винсент М., Сунь Ю., Ю Х., Ван Дж. и др. (2005). Полная последовательность генома бактериофага Т5. Вирусология 332, 45–65. 10.1016/j.virol.2004.10.049 [PubMed] [CrossRef] [Google Scholar]
- Wiles S., Clare S., Harker J., Huett A., Young D., Dougan G., et al. (2005). Органоспецифичность, динамика колонизации и клиренса in vivo после перорального заражения мышиным патогеном Citrobacter rodentium . Клетка. микробиол. 7, 459–459. 10.1111/j.1462-5822.2004.00479.x [PubMed] [CrossRef] [Google Scholar]
Статьи из Frontiers in Microbiology предоставлены здесь с разрешения Frontiers Media SA
Портативный твердотельный накопитель Samsung T5 | Спецификации и функции
Мы используем файлы cookie, чтобы улучшить ваше взаимодействие с нашим веб-сайтом, а также показать вам, как вы работаете на нашем веб-сайте, и показать вам релевантную рекламу.
Управляйте настройками наших файлов cookie ниже.Основные файлы cookie
Эти файлы cookie необходимы, поскольку они позволяют вам перемещаться по веб-сайту. Эту категорию нельзя отключить.
Компания Домен Гугл semiconductor.samsung.com, image.semiconductor.samsung.com, smetrics.samsung.com Аналитические или эксплуатационные файлы cookie
Эти файлы cookie собирают информацию о том, как вы используете наш веб-сайт, например, какие страницы вы посещаете чаще всего. Вся информация, которую собирают эти файлы cookie, используется для улучшения работы веб-сайта.
Аналитические или эксплуатационные файлы cookie Печенье Домен Google ajax.googleapis.com, apis. google.com, calendar.google.com, developer.google.com, docs.google.com, google.com, maps.googleapis.com, электронные таблицы.google.com, www.google .com, www.google.ieГугл www.google-analytics.com, www.googletagmanager.com, www.gstatic.com Adobe assets.adobedtm.com Функциональные файлы cookie
Эти файлы cookie позволяют нашему веб-сайту запоминать сделанный вами выбор (например, ваше имя пользователя, язык или регион, в котором вы находитесь) и адаптировать веб-сайт для предоставления вам расширенных функций и контента.
Функциональные файлы cookie Печенье Домен Назначение Акамай 176-34-86-175_s-23-203-249-81_ts-1604430438-clienttons-s.akamaihd.net, 176-34-86-175_s-23-203-249-81_ts-1604432488-clienttons-s.akamaihd. net, 176-34-86-175_s-23-203-249-90_ts-1604428164-clienttons-s.akamaihd.net, 176-34-86-175_s-95-101-143-18_ts-1604428258-clienttons-s. akamaihd.net, 176-34-86-175_s-95-101-143-24_ts-1604428321-clienttons-s.akamaihd.net, 34-242-207-243_s-23-203-249-81_ts-1604425495-clienttons-s.akamaihd.net,
34-242-207-243_s-23-203-249-81_ts-1604425563-clienttons-s.akamaihd.net, 34-242-207-243_s-23-203-249-81_ts-1604425669-clienttons-s.akamaihd. net, 34-242-207-243_s-23-203-249-81_ts-1604427540-clienttons-s.akamaihd.net, 34-242-207-243_s-23-203-249-81_ts-1604427617-clienttons-s. akamaihd.net, 34-242-207-243_s-23-203-249-81_ts-1604427664-clienttons-s.akamaihd.net, 34-242-207-243_s-23-203-249-81_ts-1604427922-clienttons- s.akamaihd.net,
34-242-207-243_с-23-203-249-81_ts-1604439090-clienttons-s.akamaihd.net, 34-242-207-243_s-23-203-249-81_ts-1604439174-clienttons-s.akamaihd.net, 34-242-207-243_s-23-203 -249-81_ts-1604441206-clienttons-s.akamaihd.net, 34-242-207-243_s-23-203-249-81_ts-1604441267-clienttons-s. akamaihd.net, 34-242-207-243_s-23 -203-249-90_ts-1604425484-clienttons-s.akamaihd.net, 34-242-207-243_s-23-203-249-90_ts-1604425610-clienttons-s.akamaihd.net,
34-242-207-243_s-23-203-249-90_ts-1604427737-clienttons-s.akamaihd.net, 34-242-207-243_s-23-203-249-90_ts-1604427797-clienttons-s.akamaihd.net, 34-242-207-243_s-23-203-249-90_ts-1604438922-clienttons-s.akamaihd.net, 34-242-207-243_s-23-203 -249-90_ts-1604438968-clienttons-s.akamaihd.net, 34-242-207-243_s-23-203-249-90_ts-1604439033-clienttons-s.akamaihd.net, 34-242-207-243_s-23 -203-249-90_ts-1604441023-clienttons-s.akamaihd.net,
34-242-207-243_s-95-101-129-82_ts-1604425732-clienttons-s.akamaihd.net, 34-245-202-11_s-23-203-249-81_ts-1604425513-clienttons-s.akamaihd. сеть, 34-245-202-11_с-23-203-249-81_ts-1604427569-clienttons-s.akamaihd.net, 34-245-202-11_s-23-203-249-90_ts-1604425365-clienttons-s.akamaihd.net, 34-246-182-217_s-23-203 -249-81_ts-1604424915-clienttons-s.akamaihd.net, 34-246-182-217_s-23-203-249-81_ts-1604425000-clienttons-s. akamaihd.net,
34-246-182-217_s-23-203-249-81_ts-1604425155-clienttons-s.akamaihd.net, 34-246-182-217_s-23-203-249-81_ts-1604425567-clienttons-s.akamaihd. сеть, 34-246-182-217_s-23-203-249-81_ts-1604427446-clienttons-s.akamaihd.net, 34-246-182-217_s-23-203-249-81_ts-1604429495-clienttons-s.akamaihd.net, 34-246-182-217_s-23-203-249-90_ts-1604424817-clienttons-s.akamaihd.net, 34-246-182-217_s-23-203 -249-90_ts-1604424939-clienttons-s.akamaihd.net,
34-246-182-217_s-23-203-249-90_ts-1604427359-clienttons-s.akamaihd.net, 34-246-182-217_s-23-203-249-90_ts-1604429563-clienttons-s.akamaihd. net, 34-246-182-217_s-95-101-129-82_ts-1604425062-clienttons-s.akamaihd.net, 34-246-182-217_s-95-101-143-18_ts-1604429398-clienttons-s. akamaihd.net, 34-246-182-217_s-95-101-143-24_ts-1604429274-clienttons-s.akamaihd.net, 34-246-182-217_s-95-101-143-24_ts-1604429365-clienttons-s.akamaihd.net,
34-246-182-217_s-95-101-143-24_ts-1604429616-clienttons-s.akamaihd.net, 364bf52c.akstat.io, 364bf5fa.akstat.io, 364bf6cc. akstat.io, 36c3fef2.akstat.io, 54-154-186-178_s-23-203-249-81_ts-1604425586-clienttons-s.akamaihd.net, 54-154-186-178_s-23-203-249-81_ts-1604429882-clienttons-s.akamaihd. сеть, 54-154-186-178_s-23-203-249-90_ts-1604425341-clienttons-s.akamaihd.net, 54-154-186-178_s-23-203-249-90_ts-1604425577-clienttons-s.akamaihd.net,
54-154-186-178_s-23-203-249-90_ts-1604425679-clienttons-s.akamaihd.net, 54-154-186-178_s-23-203-249-90_ts-1604427498-clienttons-s.akamaihd. net, 54-154-186-178_s-23-203-249-90_ts-1604431774-clienttons-s.akamaihd.net, 54-154-186-178_s-92-123-142-66_ts-1604427735-clienttons-s. akamaihd.net, 54-246-30-86_s-23-203-249-81_ts-1604425115-clienttons-s.akamaihd.net, 54-246-30-86_s-23-203-249-81_ts-1604427273-clienttons- s.akamaihd.net,
54-246-30-86_с-23-203-249-81_ts-1604427303-clienttons-s.akamaihd.net, 54-246-30-86_s-23-203-249-81_ts-1604427359-clienttons-s.akamaihd.net, 54-246-30-86_s-23-203 -249-81_ts-1604431429-clienttons-s.akamaihd.net, 54-246-30-86_s-23-203-249-81_ts-1604431547-clienttons-s. akamaihd.net, 54-246-30-86_s-23 -203-249-81_ts-1604435637-clienttons-s.akamaihd.net, 54-246-30-86_s-23-203-249-90_ts-1604427151-clienttons-s.akamaihd.net,
54-246-30-86_s-23-203-249-90_ts-1604429503-clienttons-s.akamaihd.net, 54-246-30-86_s-23-203-249-90_ts-1604429594-clienttons-s.akamaihd.net, 54-246-30-86_s-23-203-249-90_ts-1604433473-clienttons-s.akamaihd.net, 54-246-30-86_s-23-203- 249-90_ts-1604433539-clienttons-s.akamaihd.net, 54-246-30-86_s-88-221-134-224_ts-1604435698-clienttons-s.akamaihd.net, 54-246-30-86_s-95- 101-129-96_ts-1604424926-clienttons-s.akamaihd.net,
54-246-30-86_s-95-101-129-96_ts-1604424989-clienttons-s.akamaihd.net, 54-75-39-103_s-23-203-249-81_ts-1604425265-clienttons-s.akamaihd. сеть, 54-75-39-103_s-23-203-249-81_ts-1604425415-clienttons-s.akamaihd.net, 54-75-39-103_s-23-203-249-90_ts-1604425504-clienttons-s.akamaihd.net, 54-75-39-103_s-95-101-143-24_ts-1604432234-clienttons-s.akamaihd.net, 54-75 -41-190_s-23-203-249-81_ts-1604424935-clienttons-s.akamaihd.net,
54-75-41-190_s-23-203-249-81_ts-1604425058-clienttons-s. akamaihd.net, 54-75-41-190_s-23-203-249-81_ts-1604425120-clienttons-s.akamaihd. net, 54-75-41-190_s-23-203-249-81_ts-1604425189-clienttons-s.akamaihd.net, 54-75-41-190_s-23-203-249-81_ts-1604427540-clienttons-s. akamaihd.net, 54-75-41-190_s-23-203-249-90_ts-1604424875-clienttons-s.akamaihd.net, 54-75-41-190_s-23-203-249-90_ts-1604425270-clienttons-s.akamaihd.net,
54-75-41-190_s-23-203-249-90_ts-1604427110-clienttons-s.akamaihd.net, 54-75-41-190_s-23-203-249-90_ts-1604429433-clienttons-s.akamaihd. net, 54-75-41-190_s-23-203-249-90_ts-1604429456-clienttons-s.akamaihd.net, 54-75-41-190_s-92-123-140-11_ts-1604427291-clienttons-s. akamaihd.net, 54-75-41-190_s-92-123-140-11_ts-1604427412-clienttons-s.akamaihd.net, 54-75-41-190_s-95-101-129-96_ts-1604425019-clienttons-s.akamaihd.net,
54-75-41-190_s-95-101-143-18_ts-1604429529-clienttons-s.akamaihd.net, 684dd305.akstat.io, 684dd306.akstat.io, 684dd307.akstat.io, 684dd308.akstat.io, 684dd309.akstat.io, 684dd30a.akstat.io, 684dd30c.akstat.io, 684dd30d. akstat.io, 6852bd07.akstat.io, 6852bd08.akstat.io, 6852bd09.akstat.io, 6852bd0a.akstat.io, 6852bd0b. akstat.io, 6852bd0c.akstat.io, 6852bd0d.akstat.io, 6852bd0e.akstat.io, 6852bd0f.akstat.io, 6852bd10.akstat.io, 6852bd11.akstat.io, 6852bd12.akstat.io,
6852bd13.akstat.io, 6852bd14.akstat.io, 685d5b18.akstat.io, 685d5b19.akstat.io, 685d5b1b.akstat.io, 686eb51b.akstat.io, 686eb704.akstat.io, bcsecure01-a.akamaihd.net, Brightcove04pmdo-a.akamaihd.net, ds-aksb-a.akamaihd.net, el24ucyccuqvax5bs2kq -pblhb6-a723eeea5-clientnsv4-s.akamaihd.net, el24ucyccuqvax5bt4yq-ptbmxa-6ef8e4803-clientnsv4-s.akamaihd.net, el24ucyccuqwcx5bs4uq-p03zy7-676237e5e-clientnsv4-s.akamaihd.net
el3lnwiccuqvax5bstvq-pch0tk-1cdf76638-clientnsv4-s.akamaihd.net, el3lnwiccuqvax5buy2q-pqnfkn-f673b4feb-clientnsv4-s.akamaihd.net, el3lnwiccuqvax5buzkq-pl30i3-08d7d87df-clientnsv4-s.akamaihd.net, el3lnwiccuqwcx5bu4ya-pyg66y-cb19A994E-clientNSV4-S.Kamaihd.net, el3lnwiccuqxax5bstjq-puyi2b-1f022524f-clientnsv4-s.kamaihd.net, el3lnwiccuqxax5bsuua-pioden-695058c8f-clientnsv4-s. akmahihd.-byden-695058c8f-clientnsv4-s.akmahihd.-byden-pioden-695058c8f-clientnsv4-widen-pioden-pioden-695058c8f-client-client-cle сеть,
el3lnwiccuqxax5btzpq-pbifp1-07760bdf0-clientnsv4-s.akamaihd.net, el3lnwiccuqxax5bu23q-p2ez1a-7d289db29-clientnsv4-s.akamaihd.net, el3lnwixzp4swx5bs5pq-pnfw20-03cb87b70-clientnsv4-s.akamaihd.net, el3lnwixzp4swx5bsryq-p52tb9-f3dab0dd0-clientnsv4- s.akamaihd.net, el3lnwixzp4swx5bu35q-pdannf-fd1139023-clientnsv4-s.akamaihd.net, el3lnwixzp4swx5buxna-pyccr1-f710a073b-clientnsv4-s.akamaihd.net, el3lnwky3wdkax5bsxbq-p3hn9l-a2a7437e4-clientnsv4-s.akamaihd.net,
el3lnwky3wdkex5bt23a-pfcryk-8b7c1430e-clientnsv4-s.akamaihd.net, elzm742y3wdkex5bs4lq-p0p40d-3a2e745b5-clientnsv4-s.akamaihd.net, elzm742y3wdkex5bzofa-pqb527-96b6b1fc9-clientnsv4-s.akamaihd.net, elzm74yccuqvax5b2szq-pf5z0b-8e0fe713e-clientnsv4- s.akamaihd.net, elzm74yccuqvax5bs5nq-pt4puj-60e29ce0a-clientnsv4-s.akamaihd.net, elzm74yccuqvax5bzo4a-ptxi68-223a872ab-clientnsv4-s.akamaihd.net, elzm74yccuqwcp8b4b2t0a—clientnsv4-s. akamaihd.net,
elzm74yccuqwcx5btaca-p2p13t-2edd5f4d6-clientnsv4-s.akamaihd.net, elzm74yccuqwcx5buakq-p7s1ie-7095e2510-clientnsv4-s.akamaihd.net, elzm74yccuqxax5b2o7q-partxm-0ba99e22d-clientnsv4-s.akamaihd.net, elzm74yccuqxax5bs6fa-pnivpg-c4bb-clientnsv4- S.AkamaIhd.net, Elzm74yccuqxax5bt5qq-pcrjf9-bdc24fa26-clientnsv4-s.kamaihd.net, elzm74yccuqxax5bzp4q-pkl6rx-fb475a90e-clientnsv4-s.alcamahd.net, elzm7475a90e-clientnsv4-s.kamahihd.nel, elzm7475a90e-clientnsv4-s.kamamahd.nel6475a90e-clientnsv4-s.kamamaihd.-pient-pient-pient-s-p
elzm74yxzp4swx5bs7cq-p4s4el-cd1a19887-clientnsv4-s.akamaihd.net, elzm74yxzp4swx5bt4ka-p0qvim-2e8a5e71e-clientnsv4-s.akamaihd.net, elzm74yxzp4swx5bt6hq-pzy1yp-35d9d01e0-clientnsv4-s.akamaihd.net, elzm74yxzp4swx5bt7mq-p1duy0-1060998fa-clientnsv4-s.akamaihd. net, elzm74yxzp4swx5bucja-p0twy9-19851792c-clientnsv4-s.akamaihd.net, elzm74yxzp4swx5bzqza-pn76ir-1c0c55ff7-clientnsv4-s.akamaihd.net, elzm74yxzp4swx5bzsda-pqodge-888ec876f-clientnsv4-s.akamaihd.net,
g2nlvmqcchiscx5bva5a-pwotro-14b66ca5a-clientnsv4-s. akamaihd.net, g2nlvmqccuqvax5bs7hq-p4vzcl-ad59a5fd9-clientnsv4-s.akamaihd.net, g2nlvmqccuqxax5bsz6q-pm3a6a-3feb7d021-clientnsv4-s.akamaihd.net, g2nlvmqxzp4swx5bs5uq-pd12b9-62c8cb38d-clientnsv4-s.akamaihd.net, g2nlvmqxzp4swx5bt3va-p7puv0-d4fafcfea-clientnsv4-s.akamaihd. сеть, g2nlvmsy3wdkax5bs5zq-p675cj-d0b1fd299-clientnsv4-s.akamaihd.net, g33b4vqccuqvax5btwhq-pfp8ei-5c0ea4329-clientnsv4-s.akamaihd.net,
g33b4vqccuqvax5btytq-pupet4-0083df35c-clientnsv4-s.akamaihd.net, g33b4vqccuqvax5btzpq-pr1f2f-01d5fb765-clientnsv4-s.akamaihd.net, g33b4vqccuqvax5bvzcq-phk9tj-828709858-clientnsv4-s.akamaihd.net, g33b4vqccuqvax5bw2bq-py1x2v-a7310f6e5-clientnsv4-s.akamaihd.net, g33b4vqccuqvax5bx3za-pge3ox-a91a32353-clientnsv4-s.akamaihd.net, g33b4vqccuqwcx5bu4na-pqdvvi-3aaa5c611-clientnsv4-s. akamaihd.net, g33b4vqccuqwcx5bwzaq-pvw5k6-d3e3dcd05-clientnsv4-s.akamaihd.net,
g33b4vqccuqwcx5bx22q-p8kovq-e038e0c0c-clientnsv4-s.akamaihd.net, g33b4vqccuqxax5bstpa-p4rsfx-bd0382a30-clientnsv4-s.akamaihd.net, g33b4vqccuqxax5btyeq-poz8cc-9955b8a36-clientnsv4-s. akamaihd.net, g33b4vqxzp4swx5bu27q-pxv1vf-89db7a111-clientnsv4-s.akamaihd.net, g33b4vqxzp4swx5bv25q-pt8447-731cc407d-clientnsv4-s.akamaihd.net, g33b4vsy3wdkax5bswnq-plqmrf-ff7289811-clientnsv4-s.akamaihd.net, g33b4vsy3wdkex5bsuoq-p56ka1-9bf23f300-clientnsv4-s.akamaihd. сеть,
gzfsozyccuqwcx5bs3dq-p2yzo8-69eb1f4d7-clientnsv4-s.akamaihd.net, gzfsozyccuqwcx5bs4qa-p299q7-a9521f4ee-clientnsv4-s.akamaihd.net, gzfsozyccuqwcx5bsyyq-pv69oz-aed1b09c6-clientnsv4-s.akamaihd.net, gzfsozyccuqwcx5bwfuq-pw4gfb-c2c42381f-clientnsv4- S.AkamaIhd.net, GZFSTPQCCUQVAX5BSSVQ-P0X8HM-7A3D7367F-CLIENTNSV4-S.KamaIHD.NET, GZFSTPQCCUQVAX5BSXSQ-P2OAJS-B2E67F00B-CLIENSV4-S.AkamaIHD.NET, GZFSTPS-B2E67F00B-CLIentNSV4-S.KamaIHD.NET, GZFST-PQUN-P-SHINBAX-P-SHINBAX-P-SHINBAX-P-SHINBAX-P-SHINBAX-P-57
gzfstpqccuqvax5bu2ia-p6uwyn-30e7a92df-clientnsv4-s.akamaihd.net, gzfstpqccuqwcx5bswqa-pplxq4-ee58ceb89-clientnsv4-s.akamaihd.net, gzfstpqccuqwcx5bu3mq-p6qff7-f4c4075e7-clientnsv4-s.akamaihd.net, gzfstpqccuqwcx5buz4q-pbk4m8-d20c90e54-clientnsv4-s. akamaihd. сеть, gzfstpqccuqxax5bt4ka-p3fi1s-1fcad7cd5-clientnsv4-s.akamaihd.net, gzfstpqxzp4swx5bsttq-p683qt-2c3f6e21e-clientnsv4-s.akamaihd.net, gzfstpqxzp4swx5bsu5q-damaihd.sakientf-3b5424
gzfstpqxzp4swx5bsvra-ps8whv-800c4ca06-clientnsv4-s.akamaihd.net, gzfstpqxzp4swx5bt2ka-p3owfu-9bef421db-clientnsv4-s.akamaihd.net, gzfstpqxzp4swx5btynq-p80cg4-5fbda6ae3-clientnsv4-s.akamaihd.net, gzfstpsy3wdkax5btvta-pc4hb3-c24fbde0b-clientnsv4-s.akamaihd.s.net, i03fd9f40 i03fa4400-ds-aksb-a.akamaihd.net, i03faac00-ds-aksb-a.akamaihd.net, i03fae300-ds-aksb-a.akamaihd.net, i03fb4f00-ds-aksb-a.akamaihd.net, i22f29600- ds-aksb-a.akamaihd.net,
i22f44600-ds-aksb-a.akamaihd.net, i22f47c00-ds-aksb-a.akamaihd.net, i22f4a100-ds-aksb-a.akamaihd.net, i22f55c00-ds-aksb-a.akamaihd.net, i22f5ca00- ds-aksb-a.akamaihd.net, i22f6b100-ds-aksb-a.akamaihd.net, i22f7bf00-ds-aksb-a.akamaihd.net, i22fdb800-ds-aksb-a.akamaihd.net, i22fdd700-ds- aksb-a.akamaihd.net, i3430c200-ds-aksb-a.akamaihd.net, i34d04400-ds-aksb-a.akamaihd. net, i34d71700-ds-aksb-a.akamaihd.net, i36486200-ds-aksb- a.akamaihd.net, i364b2700-ds-aksb-a.akamaihd.net,
i369aba00-ds-aksb-a.akamaihd.net, i36d85900-ds-aksb-a.akamaihd.net, i36d86800-ds-aksb-a.akamaihd.net, i36e56b00-ds-aksb-a.akamaihd.net, i36f61e00- ds-aksb-a.akamaihd.net, i36f6c000-ds-aksb-a.akamaihd.net, i3f23f800-ds-aksb-a.akamaihd.net, ib0225600-ds-aksb-a.akamaihd.net, s.go- mpulse.net, пробная-eum-clientnsv4-s.akamaihd.net, пробная-eum-clienttons-s.akamaihd.net, warfnl2y3wdkex5buhra-pvnsej-42dd2535b-clientnsv4-s.akamaihd.net,
warfnlyccuqvax5bvjta-pivu9l-324052216-clientnsv4-s.akamaihd.net, warfnlyccuqxax5bwjua-pt5xj8-63e5f59c4-clientnsv4-s.akamaihd.net, warfnlyxzp4swx5bugca-p9ihiy-2a56daf9f-clientnsv4-s.akamaihd.net, warfnlyxzp4swx5buiqq-p5eemy-20706e9d7-clientnsv4-s.akamaihd.net Для обеспечения оптимизированного качества изображения и повышения скорости загрузки страницыДля обеспечения оптимального качества изображения и увеличения скорости загрузки страницы Amazon
(Облачный фронт)Amazon (облачный фронт) d15mv1adrb1s6e. cloudfront.net, d1vp9jkpfdwr15.cloudfront.net, d25jv1xpupcva6.cloudfront.net, d2cmqkwo8rxlr9Для ускорения например, изображения, таблицы стилей, JavaScript и т. д.) для зрителей по всему мируЧтобы ускорить доставку вашего статического контента (например, изображений, таблиц стилей, JavaScript и т. д.) зрителям по всему миру Брайтков admin.brightcove.com, metrics.brightcove.com, player.brightcove.net, sadmin.brightcove.com, vjs.zencdn.net Для поддержки потокового видео Рекламные файлы cookie
Эти файлы cookie собирают информацию о привычках вашего браузера. Они помнят, что вы посещали наш веб-сайт, и делятся этой информацией с другими организациями, такими как рекламодатели.
рекламные файлы cookie Печенье Домен Facebook atdmt.com, connect.facebook. net, cx.atdmt.com, facebook.com, www.facebook.comРеклама Google ad.doubleclick.net, adservice.google.com, adservice.google.ie, cm.g.doubleclick.net, doubleclick.net, googleads.g.doubleclick.net, pubads.g.doubleclick.net, static.doubleclick .net, stats.g.doubleclick.net, www.googleadservices.com Гугл s.ytimg.com, www.youtube.com, youtube.com Linkedin ads.linkedin.com, linkedin.com, px.ads.linkedin.com, www.linkedin.com Технические характеристики Master T5 T5 Circular fluorescent 8711500642516
Характеристики лампы
- Диммируемый
Да
- Розетка
2GX13
- Технология
Флуоресцентный
Размеры лампы
- Высота
305 мм
- Ширина
18 мм
Прочность
- Номинальный срок службы
12 000 часы)
Световые характеристики
- Применение
Дополнительная яркость
- Индекс цветопередачи (CRI)
81
- Цветовая температура
4000 К
- Категория цвета света
Холодный белый
- Номинальный световой поток
4210 просвет
Потребляемая мощность
- Мощность
55,7 Вт
- Этикетка энергоэффективности (EEL)
г
Технические характеристики
- Световой поток
4 200 просвет
- Форма
Круглый флуоресцентный
- Координата цветности Y
0,38
- Координата цветности X
0,38
Размеры и вес упаковки
- EAN/UPC — продукт
8711500642516
- Вес нетто
0,110 кг
- Вес брутто
0,230 кг
- Высота
32.
300
см- Длина
30.600 см
- Ширина
2.
500
см- Номер материала (12NC)
5784013
6063 Алюминий: знакомство с его свойствами и применением
Выбор сплава играет важную роль в вашем проекте экструзии алюминия.
6063 алюминий является наиболее популярным сплавом для экструзии, поэтому его следует учитывать. Этот сплав широко используется в архитектуре. Из-за этого его часто называют архитектурным алюминием.
Это сплав средней прочности, который используется для таких применений, как перила, оконные рамы, дверные рамы, крыши, балюстрады, рамы вывесок, торговое оборудование, ирригационные трубы, строительные изделия, электрические, морские, трубопроводы, оборудование для отдыха, хранение цистерны, рамы грузовиков и прицепы.
Этот сплав часто экструдируют в трубы и каналы для использования в архитектурных проектах.
Вот некоторые общие характеристики 6063
Этот сплав обладает средней прочностью, хорошей коррозионной стойкостью, свариваемостью и обрабатываемостью. Он также достаточно поддается механической обработке. Загрузите PDF-файл с его характеристиками и свойствами материала.
Характеристика Оценка Прочность Средняя Коррозионная стойкость GOOD СВОСА И ВОЗДАТЕЛЬНОСТЬ GOOD . достигнуто.Свойства материалов 6063-T5, 6063-T6, 6063-T832 Алюминий
Одними из наиболее распространенных вариантов отпуска для этого сплава являются T5, T6 и T832.
Физические свойства алюминия 6063
Здесь мы можем увидеть плотность сплава. Обратите внимание, что нет разницы в плотности между состояниями Т5, Т6 и Т832.
СВОБОДА 6063-T5 6063-T6 6063-T832 DENTION 9000 9000 2,70557 DENSION 9000 9000 2.70557 9000 9000 2,708 2.70557 DENSIT 0,0975 фунта/дюйм³ 2,70 г/куб.см | 0,0975 фунта/дюйм³ 2,70 г/куб.см | 0,0975 фунта/дюйм³ Механические свойства алюминия 6063
Здесь мы можем увидеть предел прочности при растяжении, предел текучести и модуль упругости для состояний Т4 и Т6.
Собственность 6063-T5 6063-T6 6063-T832 TENSILE. 27000 psi241 МПа | 35000 psi 290 МПа | 42000 psi Предел текучести 145 МПа | 21000 фунтов на квадратный дюйм 214 МПа | 31000 psi 269 МПа | 39000 psi Модуль упругости 68,9 ГПа | 10000 тыс.фунтов/кв.дюйм 68,9 ГПа | 10000 тыс.фунтов/кв.дюйм 69,0 ГПа | 10000 ksi Тепловые свойства алюминия 6063
Здесь мы можем увидеть некоторые тепловые свойства сплава, включая коэффициент теплового расширения и теплопроводность. Включены состояния Т5, Т6 и Т832.
Property 6063-T5 6063-T6 6063-T832 Coefficient of Thermal Expansion @ 20.0 — 100 °C Temp 21.8 μm/m-°C | 12,1 мкдюйм/дюйм-°F 21,8 мкм/м-°C | 12,1 мкдюйм/дюйм-°F 21,8 мкм/м-°C | 12,1 мкдюйм/дюйм-°F Теплопроводность 209 Вт/м-K | 1450 БТЕ-дюйм/час-фут²-°F 200 Вт/м-K | 1390 БТЕ-дюйм/час-фут²-°F 200 Вт/м-K | 1390 БТЕ-дюйм/час-фут²-°F Примечание.
Приведенные выше свойства материала взяты из Matweb, здесь, здесь и здесь. Обратите внимание, что они не могут быть проверены на 100%.Возможно, вас интересует состав сплава 6063.
Химический состав алюминия 6063
Этот сплав является частью серии сплавов 6000. Таким образом, его основными легирующими элементами являются магний и кремний. Магний добавляют для повышения прочности, а кремний добавляют для снижения температуры плавления металла. Здесь мы можем увидеть рекомендации по химическому составу алюминия 6063.
9003
005Element Minimum % Maximum % Magnesium 0.45 0.9 Silicon 0.2 0.6 Iron No Min 0,35 Медь Нет Мин 0,1 Марганец Нет Мин 0,1 No Min 0. 1Zinc No Min 0.1 Titanium 0.0 0.1 Other Elements No Min 0.05 each0.15 in total Теперь, когда мы увидели химический состав 6063, давайте поговорим о том, как этот сплав сравнивается с некоторыми другими ключевыми вариантами.
Сравнение алюминия 6063 с другими сплавами
Сплав 6063 является наиболее распространенным для экструзии, но вы можете рассмотреть другие варианты для вашего конкретного применения. Ниже приведены некоторые другие распространенные сплавы, которые часто рассматриваются в качестве альтернативы.
6063 по сравнению с 6061
Эти два сплава являются наиболее популярными для экструзии: 6063 занимает первое место, а алюминий 6061 — второе место. Оба являются частью серии 6000 и в основном легированы магнием и кремнием. Таким образом, они обладают многими одинаковыми качествами.
Они обладают аналогичной коррозионной стойкостью, свариваемостью и обрабатываемостью. Они оба могут использоваться в самых разных приложениях.Одно ключевое отличие состоит в том, что 6061 является более прочным из двух, и в состоянии Т6 его прочность примерно равна прочности конструкционной стали. Таким образом, он часто используется для структурных приложений. С другой стороны, одним из преимуществ 6063 является то, что из него легче формировать сложные формы с гладкими поверхностями. Он также очень подходит для анодирования и обладает лучшей коррозионной стойкостью Узнать больше »
Резюме
6063 Алюминий является наиболее распространенным выбором для экструзии благодаря его превосходной экструдируемости, высокой коррозионной стойкости и способности формовать сложные формы. Он часто используется для архитектурных приложений из-за его приятного внешнего вида и приемлемой прочности.
Загрузите наш PDF-файл о свойствах алюминия 6063, чтобы сохранить его в качестве справочного материала.
CORE-T5 – Таблетка без компромиссов
Ядро-Т5
Terminé les compromis
Смотреть видео
Core-T5
Могущество
Коннективите
Сопротивление
Автономия
Часть с вами!Способность повышать эффективность и производительность вашего соединения
Благодарность за превосходную реактивацию, поддержка CORE-T5 без особых трудностей при использовании приложений, требующих дополнительных усилий, 8-дюймовых тандисов, используемых для управления 8-дюймовым экраном, позволяет легко управлять навигацией. -Т5.
C’est simple, cette tablete est le parfait outil de pilotage de vos activités professionalnelles sur le terrain.
CORE-T5, прочное и надежное решение Чтобы обеспечить долговечность и эффективность, CORE-T5 сертифицирован Android Enterprise Recommended. Ainsi, vous êtes assures d’une mise à jour vers le prochain OS d’Android (Android 11 vers Android 12) ainsi que la mise à jour régulière des patches de sécurité pour les les prochaines 3 années.
Android 11 и основная версия Android 12
Фаза с развитием приложений
Основные патчи безопасности
Залейте 3 таблетки и удалите их
14 Навигация и работа на всех этапах плавного перехода к чтению лекций и отзывов о CORE-T5 (2) с использованием процессора Qualcomm® Snapdragon™ 665 8-Cours.
3 x
плюс мощный процессор CORE-T4(1)
De la 4G+
où que vous soyezPassez des Appels visio, échangez des données en direct, accédez aux Applis Professionalnelles, effectuez et Transmettez des Diagnostics en temps réel tout en conservant votre mobilité.
4G+, la CORE-T5 предлагает дебет jusqu’à 10 fois plus quicke que la 4G. Elle dispose d’une big panoplie de bandes qui lui permettent d’être utilisée sur les réseaux 4G public et les réseaux 4G privés.
Диапазоны частот 4G:
- 1,2,3,5,7,8,20, 28 и 32, 2-CA, 38, 40,
- 28-38-40 : bandes dédiées aux réseaux privés
- 1,2,3, 5,7,8,20, 32, 2-CA: Bandes dédiées aux réseaux publics avec la meilleure connexion.
Optimisee pour le WI-FI d’Entreprise
Ne perdez jamais votre connexion. Core-T5 предлагает оптимизировать процесс использования корпоративных сетей Wi-Fi. Стандарты Wi-Fi 802.11 k, r, v, u, постоянное изменение без изменения, а также плавные переходы между точками доступа к мему, которые можно использовать.
Коммюнике о насыщении резерва ресурсов средствами вытеснения ресурсов и определения приоритетов потоков CORE-T5.
Поддерживает комплекс протоколов миссий, анализирует интеграцию с модемом Qualcomm (поддержка функций Release 12 3GPP) и включает в себя частные решения для использования PMR (Radio Mobile Professionnelle) и других групп 3 8, 0, 0
.
.. En salle de commandement
comme au plus pres du terrainОрганизует и проводит конференции и группы по обсуждению вопросов, связанных с приложениями, а также с привлечением специалистов по реализации миссий для CORE-T5.
Appuyez
et parlez.Организация групп по проведению конференций и координация оборудования по временной программе X-TALK для оборудования CROSSCALL, которое трансформирует ваш планшет в настоящую рацию
Бутоны №
программируемых,
восточных использований.Que vous soyez droitier ou gaucher, à vous de définir les raccourcis que vous souhaitez. CORE-T5 предлагает дизайн двух кнопок, программируемых устройств, совместимых с приложениями Push To Talk (PTT) и защитными функциями Isolé (PTI), а также другими функциями планшета (факел, фотография устройства и т. д.).
2 программируемые кнопки Параметры приложения де вотре выбор.
Служба спасения
vos raccourcisConservez un accès quicke aux fonctions de la tablette.
Установка и рекогносцировка программируемых бутонов на основе исследований, посвященных тому, что у вас есть возможности, и быстрому и удобному доступу к raccourcis grâce à leur texture et leur taille différentes par rapport aux autres boutons.Место для бутоньерок Évitez les appuis par inadvertance
Gagnez en Rapidité Удобный доступ к программируемым кнопкам
Des boutons faits pour
supporter la pressionИспользуемые таблетки, используемые в качестве средства для бордирования, смешиваются с морской водой, хлором, муссом, соболем и микрочастицами, содержащими микрочастицы. Aussi Durables que la CORE-T5, les boutons programmables conservent leur étanchéité même après 400 000 нажатий, soit plus de 200 pressions par jour sur 5 ans. Certifée IP68, la CORE-T5 peut encaisser une immersion jusqu’à 2m de profondeur подвесной 30 минут.
Этаншейте прочный
400 000 нажатий подвеска на 200 нажатий в день 5
Сертификация IP68 2 м — 30 мин
UNE TABLETTE PARÉE
POUR LE LONG-TERMESoyez serein et pour longtemps.
Garantie 5 ans et pensée dans les exigences CROSSCALL en termes de durabilité, la
CORE-T5 гарантирует, что aux Professionalsnels les plus exigeants qu’elle est parfaitement адаптируется к leur quotidien ainsi que leurs обычаи.Навык плюс
Ла fabilité ? C’est par ici.
Американская военная норма MIL STD –810H
Quel que soit votre quotidien, la CORE-T5 estadatée. En effet, la tablette a passé haut la main les 15 test de la norme militaire MIL-STD-810H.
Сопротивление на высоте Atteignez les sommets sans crainte.
Радиационный солер N’ayez plus peur des coups de chaud.
Бруйяр Салин Используйте свой смартфон в морской среде без рискованного окисления.
Термический шоколад Passez du chaud au froid et inversement en quelques secondes.
Загрязнение жидкостями Сопротивление дизельному топливу
Запас холодный Запас в окружающей среде при -50°.
Складской шкаф Stocké dans un environnement jusqu’à +71°.
Шоколад Votre téléphone ne craint pas les chutes.
Вибрация Votre téléphone encaisse ле secousses и др ле вибраций.
Выносливый Подвеска Humidité et chaleur 28 дней.
Ускорение Votre téléphone encaisse sans broncher l’accelération soudaine (лоток).
Плюи Ваш смартфон не craint pas les averses.
Холодный интенсивный Utilisez votre téléphone dans les régions les plus froides.
Шалер интенсивный Остальная функция à +71°.
Плюэ вергласанте Сопротивление jusqu’a 13 мм от геля, и остальная часть foctionnel après le dégel.
Функциональный и устойчивый экран
Функциональный и устойчивый экран
CORE-T5 с покрытием из стекла Corning Gorilla® Glass 3 толщиной 1,1 мм с защитой от лучей и шоколада в дизайне davantage l’écran. Pratique, vous pouvez l’utiliser avec des gants ou encore les doigts mouillés grâce aux fonctions GLOVE et WET TOUCH.
Стекло Corning Gorilla® Glass 3 толщиной 1,1 мм Защита от шока и лучей
Прикосновение к перчатке Можно использовать с брюками jusqu’à 4mm d’épaisseur
Wet touch Можно использовать avec les doigts mouillés
Votre environnement, Son Terrain de Jeu
Vous n’aurez plus peur des chutes et ça, ça change tout.
Dans le cadre de son développement, la CORE-T5 a subi et encaissé des tests de chute sur les 6 faces de la tablette à 1,5 mètres de hauteur et directement sur une dalle de beton, la matière la plus éprouvante.Тестовые желоба 6 сторон 1,5 м высоко над бетоном
сила
50 часов связь
22 часа en навигация GPS
26ч d’utilization d’applications métiers.
7000 мАч Емкость батареи
UNE TABLETTE
SUR-MESUREОптимальная премия
Оптимальная премия Profitez d’une. Выпускается с экраном 8 дюймов и форматом 227,6 x 135 x 13,8 мм, весит 594 г, CORE-T5 обладает размерами парфелей для обеспечения лучших компромиссов между хвостами, дозировкой и удобством использования.
Экран 8’’
Комфорт использования
Идеальный формат
Оптимальная цена
Свободные сети
Свободные сети для защиты ваших планшетов. Оборудование для технологии X-LINK™ и X-BLOCKER, а также для CORE-T5 является основой системы аксессуаров CROSSCALL.
Для подключения к X-TACTICAL, для поддержки тактических жилетов с MOLLE, для X-CHEST или для бандульера X-STRAP, для подключения к CORE-T5 через порт сети оставшиеся свободные движения.
Аксессуары, адаптирующиеся ко всем профилям
X-TACTICAL, X-CHEST, X-STRAP
Le petit rien qui fait tout
Pour davantage de sérénité dans chacun de vos Mouvements, la CORE-T5 est livrée avec un sécurité plugable à l’encoche directement intégrée dans son design.
х-блокатор
Avec le X-BLOCKER,
le renfort mécaniquePret à booster les possibilités et les использование фиксации X-LINK™? Благодарите за устойчивость к силе падения 20 кг, X-BLOCKER vous permet de sécuriser la fixation votre téléphone lors de vos meilleurs descentes avec le X-BIKE ou encore vos plus belles blurs avec le X-CHEST.
Фиксез
Фиксация и ориентация в простом портретном режиме / портрет
Чаргез
Быстрая и эффективная перезарядка, 98% мощности передачи
Трансферес
Простое использование.
About the author